Přehled gamet a oplodnění
Pohlavní rozmnožování se týká pohlavních zárodečných buněk (gamet), které produkují rodiče, kombinací oboupohlavních zárodečných buněk (např. spermie a vaječné buňky) se stávají oplodněným vajíčkem a z oplodněného vajíčka se pak vyvíjí nový jedinec, tzv. pohlavní rozmnožování. Rozsáhlá variabilita kombinací genů při pohlavním rozmnožování může zvýšit schopnost potomků přizpůsobit se přírodnímu výběru. Náhodně kombinované geny v potomstvu při pohlavním rozmnožování mohou, ale nemusí být pro druh prospěšné, přinejmenším však zvyšují šance několika jedinců na přežití v nepředvídatelném a neustále se měnícím prostředí, a tím prospívají druhu. Pohlavní rozmnožování může také podporovat šíření příznivých mutací v populaci. Pokud mají dva jedinci druhu příznivé mutace na různých místech, budou v populaci při nepohlavním rozmnožování oba mutanti soupeřit až do jedné eliminace, není možné zachovat obě příznivé mutace najednou. V populacích s pohlavní reprodukcí se však díky páření a rekombinaci mohou tyto dvě příznivé mutace současně dostat do genomu téhož jedince a současně se šířit v populaci. Z výše uvedených důvodů pohlavní rozmnožování urychluje proces evoluce. Za více než 3 miliardy let biologické evoluce na Zemi zůstaly první dvě miliardy let života ve fázi nepohlavního rozmnožování a evoluce probíhala pomalu, od poslední 1 miliardy let se tempo evoluce výrazně zrychlilo. Kromě změn v zemském prostředí (jako je vznik atmosféry obsahující kyslík atd.) je hlavní příčinou také vznik a rozvoj pohlavního rozmnožování.
Gametami
Gametami se rozumí zralé buňky produkované reprodukčním systémem při pohlavním rozmnožování organismu, označované jako pohlavní buňky. Gamety se dělí na samčí gamety a samičí gamety. Samičí gamety živočichů a rostlin se obvykle nazývají vaječné buňky a samčí gamety se nazývají spermie. Spermie jsou poměrně malé, ale mohou se pohybovat a vstupovat do vaječné buňky ve tvaru štíra. Vajíčková buňka je poměrně velká a neplavecká. Například objem vaječné buňky mořského ježka je 10 000krát větší než objem spermatické buňky. I když se samčí a samičí gamety liší objemem, jaderná DNA, kterou poskytují potomkům, je stejná, to znamená, že každá poskytuje sadu genomů. Vzhledem k velké velikosti vaječných buněk však cytoplazmatickou strukturu a cytoplazmatickou DNA dceřiných buněk poskytují v podstatě vaječné buňky. Gamety hrají důležitou roli v biologických výpočtech. Prostřednictvím genetických map lze jasně pozorovat tok genů a strukturu genotypů potomstva. Počet gamet v první generaci hybridů a relativních znaků u hybridů je 2n-tá mocnina (n je logaritmus relativních znaků). Když živé tělo tvoří gamety, párové genetické faktory se od sebe oddělují a vstupují do různých gamet. Gamety obsahují pouze jeden z každého páru genetických faktorů. Při oplození je kombinace samčí a samičí gamety náhodná. Vzhledem k rodu gamet vzniklých meiózou, různorodosti složení chromozomů, rozdílnosti genetického materiálu různých gamet a náhodnosti kombinace vaječných buněk a spermií v procesu oplození musí být potomci téhož rodiče různorodí.
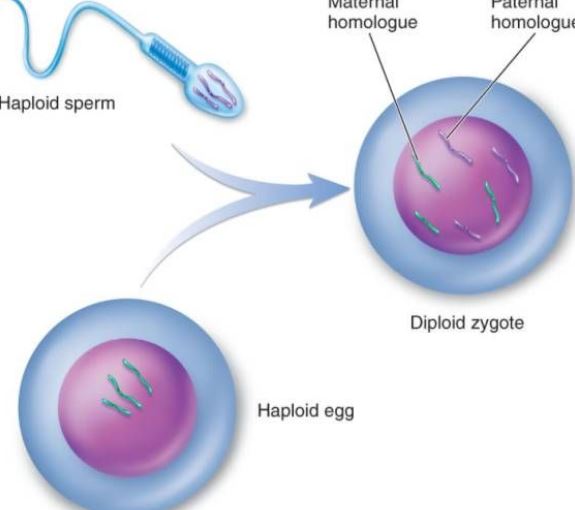
Oplodnění
Oplodnění je proces, při kterém se vajíčka a spermie spojí v jednu zygotu. Je základním rysem pohlavního rozmnožování a je všudypřítomný v živočišném i rostlinném světě, ale často se o něm hovoří jako o nejčastěji označovaném. Oplození živočichů na buněčné úrovni zahrnuje proces oplození tři hlavní fáze: aktivaci vajíčka, regulaci a amfoterní pronukleární fúzi. Aktivaci lze považovat za výchozí bod ontogeneze, který se projevuje především změnami v propustnosti výtrusné membrány, eklezií kortikálních granulí, tvorbou oplozené membrány atd. Po aktivaci dochází k úpravě, která je prvním krokem k zajištění normálního dělení oplozených vajíček. Změna prokaryotického jádra zajišťuje dědičnost po rodičích a obnovuje diploiditu. Oplodnění nejen iniciuje replikaci DNA, ale také aktivuje genetickou informaci, jako je mRNA a rRNA, ve vajíčku, aby se syntetizovaly bílkoviny potřebné pro vývoj embrya. Po 6 až 7 dnech po oplodnění blastocysta zona pellucida zaniká a postupně se pohřbívá a je pokryta endometriem, což se nazývá implantace oplodněného vajíčka. Spermie živočicha nemají zjevnou chemotaxi jako spermie nižších rostlin, např. mechorostů, ale do blízkosti vajíčka se dostávají spíše aktivním pohybem nebo ciliárním pohybem buněk pohlavního epitelu. Kapacitace spermií: Je známo, že při průchodu mnoha savčích spermií samičím pohlavním ústrojím nebo kumulusem se odstraní cizí bílkovina, která spermii obaluje, a změní se fyzikální a biologické vlastnosti plazmatické membrány spermie, což spermii umožní získat energii a účastnit se procesu oplození. Když se kapacitní spermie savců dotkne membrány vajíčka nebo zona pellucida vajíčka, specificky se naváže na glykoprotein na membráně vajíčka, který stimuluje spermii k produkci energie. Akrozomová reakce pomáhá spermii dále překonat membránu vajíčka. Na vajíčku mořského ježka je akrozomální reakcí spermie určitá polysacharidová látka v peri-membránovém filmu. Většina vajíček má na periferii vaječné membrány a tloušťka různých vaječných membrán je různá. Hlavní složkou je mucin nebo mukopolysacharid; jen některá vajíčka jsou holá, například vajíčka střevlíkovitých. Akrozomová reakce: Když spermie projde membránou vajíčka, dochází k procesu spojování. Ten probíhá volně, bez zásahu vnější teploty a nemá žádnou specifičnost. Během období adheze se protoplastový protein na akrozomální membráně mění na akrozomální protein a akrozomální protein urychluje přechod spermie přes vaječnou membránu; Je to silné spojení, může být narušeno nízkou teplotou a má specifitu. Na plazmatické membráně spermie mořského ježka byl izolován protein, který se specificky váže na glykoprotein vajíčka, tzv. vazebný protein, s molekulovou hmotností asi 30 000 molekul. Sekundární oocyty uprostřed druhého zracího dělení během ovulace spolu s okolní zona pellucida a radiační korunkou rychle vstupují do ampuly vejcovodu přes břišní dutinu díky kývání řasinek epitelových buněk vejcovodu a kontrakci svalové vrstvy. Pokud se vaječná buňka nesetká se spermií, začne obvykle během 12-24 hodin degenerovat. Jakmile se spermie dostane do kontaktu s vajíčkem, vajíčko samo projde řadou aktivačních změn. Na savčích vajíčkách je to charakterizováno kortikální reakcí, reakcí vaječníkové membrány a reakcí zona pellucida, které mohou blokovat multifertilizaci a stimulovat další vývoj vajíček. Ke kortikální reakci dochází v době splynutí spermatických buněk. Od místa splynutí kortikální částice praskají a obsah se vyplavuje, čímž se rozšiřuje kůra celého vajíčka. Reakce vaječné membrány je proces rekombinace obalu vajíčka a kortikálních granulí. Reakce zona pellucida je proces, při kterém dochází k výtoku kůry a zona pellucida z oplozené membrány, membrána vajíčka se oddělí od plazmatické membrány, receptor pro spermie v zona pellucida zmizí a zona pellucida ztvrdne. S vajíčkem může splynout pouze spermie, u které proběhla akrozomová reakce. Působením akrosinu projde spermie radiální korunkou a interaguje s molekulou glykoproteinu ZP3 receptoru spermie na zona pellucida, což spermii umožní uvolnit akrozomální enzym a vstoupit do perivitellina přes zona pellucida. Na začátku oplození je obal ekvatoriální části hlavičky lidské spermie v kontaktu s membránou vaječné buňky a poté jádro a cytoplazma spermie vstoupí do vajíčka. Poté, co spermie vstoupí do vajíčka, kortikální částice v mělké cytoplazmě vajíčka okamžitě uvolní svůj obsah na periferii membrány. Současně s tím, jak membrána kortikálních granulí splývá s buňkami vajíčka, se zvyšuje záporný povrchový náboj na povrchu buněk, čímž se potlačuje splynutí plazmatické membrány spermie s membránou vajíčka, což se nazývá kortikální reakce. Změna struktury zona pellucida se nazývá zona pellucida reakce. V této době se snižuje schopnost zona pellucida vázat spermie, což zabraňuje vzniku polyspermie a zajišťuje biologické vlastnosti lidské monospermie. Kromě zničení nebo inaktivace receptoru vázaného na spermie v zona pellucida může obsah kortikálních granulí také zvýšit zesíťování peptidových řetězců v zona pellucida, oslabit citlivost vůči akrozomové proteáze a zabránit průniku spermií. Přestože zona pellucida prochází několik spermií, do vaječné buňky se dostane pouze jedna spermie, která ji oplodní. V abnormální situaci se na oplodnění podílejí dvě spermie, a to při dvojitém oplodnění spermií. Dvě spermie současně vstoupí do vajíčka a vytvoří zárodky triploidních buněk, které jsou všechny potraceny nebo brzy po narození zemřou. Po vstupu spermie do vajíčka vajíčko rychle dokončí druhé zralé dělení. V této době se jádra spermie a vajíčka nazývají samčí pronukleus a samičí pronukleus. Obě pronuklea se postupně sblíží, jaderná membrána zmizí a chromozomy se spojí a vznikne diploidní oplozené vajíčko. Po splynutí spermií lze na povrchu vajíčka pozorovat mikroklky, které obklopují spermii, jež může být orientovaná; poté vaječná membrána splyne s plazmatickou membránou v zadní oblasti akrozomu spermie.
Reference
- Yanagimachi R. Germ cells and fertilization: why I studied these topics and what I learned along the path of my study. Andrologie. 2014, 2(6):787-93.
- Henshaw J M, Marshall D J, Jennions M D, et al. Local gamete competition explains sex allocation and fertilization strategies in the sea. American Naturalist. 2014, 184(2):32.
- Mori T, Igawa T. Gamete attachment process revealed in flowering plant fertilization. Plant Signaling & Behavior. 2014, 9(12):977715.
- Hédouin L, Pilon R, Puisay A. Hyposalinity stress compromises the fertilization of gametes more than survival of coral larvae. Výzkum mořského prostředí. 2015, 104:1-9.
Luo J, Mcginnis L K, Carlton C, et al. PTK2b function during fertilization of the mouse oocyte. Biochem Biophys Res Commun. 2014, 450(3):1212-1217.