ADVERTISEMENTS:
V tomto článku budeme diskutovat o Paramecium Caudatum:- 1. Habitat, habitat a kultura Paramecium Caudatum 2. Živočichové. Struktura Paramecium Caudatum 3. Pohyblivost 4. Výživa 5. Dýchání a vylučování 6. Osmoregulace 7. Chování 8. Rozmnožování 9. Aberantní chování při rozmnožování 10. Některé cytoplazmatické částice.
Obsah:
- Život, Habitat a kultura Paramecium caudatum
- Struktura Paramecium caudatum
- Lokomoce Paramecium caudatum
- Výživa u Paramecium caudatum
- Dýchání a vylučování u Paramecium caudatum
- Osmoregulace Paramecium Caudatum
- Chování Paramecium Caudatum
- Reprodukce u Paramecium Caudatum
- Aberantní chování při reprodukci u Paramecium Caudatum
- Některé cytoplazmatické částice popsané u Paramecium Caudatum
- Habit, Habitat and Culture of Paramecium Caudatum:
- Kultura paramecií:
- Struktura Paramecium caudatum:
- (i) Velikost a tvar Paramecium caudatum:
- (ii) Pelikula:
- (iii) Cilia:
- (iv) Infračervený systém:
- (v) Ústní rýha a cytopyge:
- (vi) Cytoplazma:
- (vii) Ektoplazma:
- (viii) Trichocysty:
- (ix) Neuromotorika a přidružená vlákna:
- (x) Endoplazma:
- (xi) Kontrakční vakuoly:
- (xii) Potravní vakuoly:
- Lokomoce Paramecium caudatum:
- (i) Metaboly neboli tělesné kontorze:
- (ii) Řasinkový pohyb:
- Výživa Paramecium caudatum:
- Mechanismus výživy:
- Trávení a vylučování Paramecium Caudatum:
- Dýchání a vylučování Paramecium caudatum:
- Chování Paramecium caudatum:
- (i) Reakce na kontakt (Thigmotaxis):
- (ii) Reakce na chemické látky (chemotaxe):
- (iii) Reakce na teplotu (termotaxe):
- (iv) Reakce na světlo (fototaxe):
- (v) Reakce na elektrický proud (galvanotaxe):
- (vi) Reakce na vodní proud (reotaxe):
- (vii) Reakce na gravitaci (geotaxe):
- Reprodukce u Paramecium Caudatam:
- (i) Příčné binární dělení:
- (ii) Konjugace:
- Aberantní chování při rozmnožování u Paramecium Caudatum:
- (i) Endomixe:
- (ii) Autogamie:
- (iii) Hemixis:
- (iv) Cytogamie:
- Některé cytoplazmatické částice popsané u Paramecium caudatum:
- (i) částice kappa:
- (ii) mµ, částice:
- (iii) Částice pí:
- (iv) Částice lambda:
Habit, Habitat and Culture of Paramecium Caudatum:
Paramecium caudatum (Gr., paramekes = podlouhlý; l., caudata = ocas) se běžně vyskytuje ve sladkovodních rybnících, tůních, příkopech, potocích, jezerech, nádržích a řekách. Zvláště hojně se vyskytuje ve stojatých rybnících bohatých na rozkládající se látky, v organických nálevech a v odpadních vodách. Paramecium caudatum je volně žijící organismus a tento druh je rozšířen po celém světě.
Kultura paramecií:
DOPORUČENÍ:
Vyjměte z rybníka ponořený plevel a vložte jej do sklenice s destilovanou vodou, sklenici přikryjte a nechte ji shnít; za několik dní se objeví roje paramecií. Nyní uvařte seno ve vodě, nálev slijte, přidejte několik zrnek pšenice a nechte stát, dokud se nezakalí bakteriemi.
Přenes Paramecia z první sklenice do této tekutiny, kde se rychle rozmnoží. Samotný nálev ze sena vytvoří Paramecia vykazující přítomnost cyst a byly zaznamenány cysty připomínající zrnka písku, ale neexistuje žádný důkaz, že Paramecia tvoří cysty, protože nebyly nikdy potvrzeny.
Struktura Paramecium caudatum:
(i) Velikost a tvar Paramecium caudatum:
Paramecium caudatum (obr. 20.1) je mikroskopický organismus a je viditelný pouhýma očima jako drobné protáhlé tělo.
PŘÍČINY:
Vypadá světle šedě nebo bíle, měří běžně 170 až 290 mikrometrů na délku a může dosáhnout délky až 300-350 mikrometrů. P. caudatum vypadá jako podrážka střevíčku nebo boty, proto je živočich běžně znám jako střevlík. Je čtyřikrát delší než široký a poněkud válcovitý s výrazně odlišnými konci.
Přední část pohybující se dopředu je štíhlá s tupým nebo zaobleným koncem, zatímco zadní konec je poněkud zašpičatělý nebo kuželovitý. Nejširší část organismu je těsně pod středem. Tělo živočicha je asymetrického tvaru a vykazuje dobře ohraničený orální nebo ventrální povrch a aborální nebo dorzální povrch.
(ii) Pelikula:
Tělo je pokryto tenkou, dvouvrstvou, pružnou a pevnou pelikulou z želatiny. Pelika drží tvar živočicha, ale je dostatečně pružná, aby umožňovala kontrakce. Pelika má dvojitou membránu, vnější membrána je spojitá s řasinkami a vnitřní membrána s ektoplasmou. Při větším zvětšení mikroskopu jsou na povrchu pelikuly patrné obdélníkové nebo šestihranné prohlubně.

Toto uspořádání je zachováno na dorzálním povrchu Paramecium, ale na ventrálním povrchu se hřebínky sbíhají vpředu a vzadu směrem k preorálnímu a postorálnímu otvoru. Každá šestiboká prohlubeň je perforována centrálním otvorem, kterým vystupuje jedno řasnaté tělísko. Na předním a zadním okraji šestibokých prohlubní jsou otvory trichocyst.
Elektronově mikroskopické studium pelikuly (obr. 20.2) Ehretem a Powersem (1957) odhalilo, že šestiboké prohlubně odpovídají pravidelným řadám dutin, alveolům. Všechny alveoly dohromady tvoří souvislou alveolární vrstvu, která je ohraničena vnější alveolární a vnitřní alveolární membránou.
Vnější vrstva leží v těsném kontaktu pod vnější buněčnou membránou. Pelikula tedy zahrnuje vnější buněčnou membránu, vnější alveolární membránu a vnitřní alveolární membránu.
(iii) Cilia:
Celé tělo je pokryto četnými, drobnými, vlasům podobnými výběžky zvanými řasinky. Řasinky se vyskytují v podélných řadách po celém těle, tento stav se nazývá holotrichózní, při kterém jsou řasinky na těle stejné. Řasinky mají stejnou stavbu jako bičíky, mají vnější protoplazmatický obal neboli plazmatickou membránu s devíti dvojitými podélnými vlákny v obvodovém prstenci. U některých řasinek není devět vnějších vláken párových.
PŘÍČINY:
Mají dvě centrální podélné fibrily, které jsou tenčí než vnější fibrily. Každé cilium vzniká z bazální granule nebo kinetosomu. Devět párů periferních fibril se spojuje a tvoří stěnu kinetosomu, kinetosom je tedy trubice, která je na svém dolním konci buď otevřená, nebo uzavřená, dvě centrální fibrily končí u většiny řasinek na úrovni pelikuly.
Z kinetosomu vychází tenký rhizoplast, který se nespojuje s jádrem. Řasinky mají i mnozí Metazoa, jejich stavba je stejná, jen bazální granule je jiná a má jemná vlákna nebo kořenová vlákna zasahující dolů do cytoplazmy. Řasinky se však od bičíků liší tím, že jsou obecně početnější a kratší.
Řasinky lze vhodně rozdělit na tělní neboli somatické, které se nacházejí na povrchu těla, a na ústní, které jsou spojeny s oblastí úst. Tělesné řasinky jsou stejné, ale na zadním konci jsou delší, odtud název caudatum. Řasinky jsou organely pro pohyb a sběr potravy, fungují také jako smyslové receptory a detekují podněty z vnějšího prostředí.

Struktura řasinek:
Řasinky a bičíky mají fibrilární složení. V základu má řasinka průměr asi 0,2 mikronu nebo 2000 A0, který může být až 10 mikronů nad povrchem buňky. Řasinky jsou ohraničeny jednotkovou membránou o tloušťce 90 A0, která se podobá plazmatické membráně a zůstává s ní spojitá. Ohraničený prostor řasinky obsahuje vodnatou látku známou jako matrix.
V matrix zůstává uloženo jedenáct podélných vláken neboli mikrotubulů. Z jedenácti fibril se dvě nacházejí uprostřed, zatímco zbývajících devět fibril zůstává uspořádáno periferně kolem centrálních fibril. Každá z devíti vnějších fibril má průměr 360A0 a skládá se ze dvou subfibril o průměru 180 až 250A0.
Tyto subfibrily se označují jako subfibrila A a subfibrila B.
DOPLŇKY:
Dílčí vlákno A je o něco větší než dílčí vlákno B. Dílčí vlákno A vydává z jedné své strany dva silné výběžky neboli ramena. Ramena subfibrily A všech vnějších fibril zůstávají směřovat ve směru hodinových ručiček. Dále se subfibrila A vyskytuje blíže ke středu řasinky než subfibrila B. Obě subfibrily mají společnou stěnu o tloušťce 50A0.
Dvě centrální fibrily nemají párové subfibrily jako devět periferních fibril, ale každá z nich obsahuje pouze jednu tubulu. Každá centrální fibrila má průměr asi 250A0 a je tvořena stěnou o tloušťce 60A0.
Obě centrální fibrily zůstávají odděleny prostorem o velikosti 350A0 a zůstávají obaleny společným pláštěm. Gibbnos (1967) uvedl, že plášť centrálních fibril vydává devět radiálně orientovaných článků nebo paprsků ke každé subfibrile A.

Elektronová mikroskopie s vysokým rozlišením odhalila, že každá z periferních a centrálních fibril řasinek a bičíků je složena z deseti až dvanácti vláken o tloušťce 40A0. Každé vlákno je korálkové. Každý korálek zůstává uspořádán v mřížkách o rozměrech 40 na 50A0 v rovině stěny bulbu. Tyto korálky jsou považovány za základní podjednotku struktury tubulu.
(iv) Infračervený systém:
Infraciliární systém se nachází těsně pod pelikulárními alveoly. Skládá se z kinetosomu neboli bazálního tělíska a kinetodesmy. Řasinky vznikají z kinetozomů a z každého kinetozomu vzniká jemné cytoplazmatické vlákno zvané kinetodesma, (obr. 20.2).
Pod pelikulou leží mírně vpravo, ale je spojena se všemi kinetosomy jedné podélné řady, podélný svazek několika kinetodesmat, kinetodesma každého kinetosomu zasahuje na určitou vzdálenost dopředu do vlastního svazku kinetodesmat.
Podélná řada kinetosomů s jejich kinetodesmaty tvoří podélnou jednotku zvanou kineta. Všechny kinetie neboli kinetia tvoří infraciliární systém řasinky. Kinetia leží v kůře pod pelikulou, jejich počet je u každého řasnatého tělíska poměrně konstantní.
Infraciliární soustava řídí a koordinuje pohyby řasinek a přináší vznik organel při dělení buněk, např. některé kinetie tvoří ústa. Při binárním dělení řasinek jsou kinetie příčně rozříznuty na dvě části, z nichž každá přechází do jedné dceřiné buňky, toto dělení se nazývá perikinetální.
(v) Ústní rýha a cytopyge:
Na ventrální straně je velká šikmá, mělká prohlubeň zvaná ústní rýha nebo peristom, která dává živočichovi asymetrický vzhled. Probíhá šikmo dozadu z jedné strany (obvykle zleva doprava, ale v některých případech zprava doleva) a končí kousek za středem těla. Ústní rýha ústí do krátké kuželovité nálevkovité prohlubně zvané vestibulum.
ÚSTY:
Vestibula vede přímo do pevného, oválného otvoru zvaného cytostom (ústí). Přímo od cytostomu směrem ke středu těla se táhne široký cytopharynx. Cytopharynx se pak prudce stáčí k zadní straně a přechází ve štíhlý zužující se jícen.
Takto je jícen zhruba rovnoběžný s povrchem těla Paramecia s výjimkou jeho zadního konce. Zde se jícen opět stáčí ke středu živočicha, aby vedl do tvořící se potravní vakuoly.
Cytopyge (označovaný také jako buněčný řitní otvor nebo anální skvrna či cytoprokt) leží na ventrálním povrchu těla téměř vertikálně za cytostomem nebo ústy. Přes cytopyge jsou vylučovány nestrávené částečky potravy. Ciliace cytofaryngu je velmi komplikovaná.
Gelei (1934) uvádí přítomnost čtyř řad a Lund (1941) pozoroval nejméně čtyři řady. Na levé stěně cytopharyngu se nachází struktura zvaná penniculus, která se spirálovitě stáčí přibližně o 90 stupňů, takže její zadní konec je na orálním (ventrálním) povrchu jícnu.
Podle Lunda se penniculus skládá z osmi řad řasinek uspořádaných do dvou těsně vedle sebe umístěných bloků. Podobný pás složený ze čtyř řad dlouhých řasinek, které jsou méně kompaktní než v penniculu, se označuje jako quadrulus. Spirálovitě se stáčí dolů po hřbetní stěně bukální dutiny a končí v blízkosti penniculu.
Někteří pracovníci nazývají penniculus a quadrulus nesprávně zvlněnou blánou. Quadrulus a penniculus řídí průchod potravy. Není známo, jak řasinky fungují, pravděpodobně se jejich vlákna rytmicky stahují, což způsobuje ohýbání. Gelei (1925) upozornil, že funkcí penniculu je vytlačování potravních elementů do těla.

(vi) Cytoplazma:
Cytoplazma se rozlišuje na úzkou, vnější neboli kortikální zónu zvanou ektoplazma a větší, vnitřní neboli dřeňovou oblast zvanou endoplazma.
(vii) Ektoplazma:
Ektoplazma (ektosarka nebo kůra) je trvalá část těla, nápadně ohraničená od endoplazmy. Ektoplazma tvoří pevnou, průhlednou, tenkou a hustou vnější vrstvu. Obsahuje trichocysty, řasinky a fibrilární struktury a navenek je ohraničena obalem zvaným pellika.
(viii) Trichocysty:
V ektoplazmě jsou v pravém úhlu k povrchu uloženy malé vřetenovité váčky zvané trichocysty. Malá skvrna na každém předním a zadním okraji šestiúhelníku označuje polohu trichocysty. Jsou vyplněny lámavou, hustou tekutinou s bobtnavou látkou, na vnějším konci je kuželovitá hlavička nebo hrot.
Trichocysty leží kolmo v ektoplazmě, otevírají se malými póry na hřebenech hexagonálních ploch pelikuly.
Vznikají z kinetozomů řasinek, pak migrují a umísťují se ve stejné vzdálenosti v endoplazmě. Když je živočich podrážděn, trichocysty se uvolňují v podobě dlouhých lepkavých vláken. Vypuštěná trichocysta má neprůhledný hrot podobný obrácenému hřebíku a dlouhý pruhovaný dřík, který však v nevypuštěném stavu není vidět a pravděpodobně vzniká až během vypouštění.
Funkce trichocyst je nejistá, ale jsou vypouštěny jako reakce na místní kontakty a poranění, mohou sloužit jako obranné organely.
To je však nejisté, protože trichocysty jsou neúčinné proti Didiniu, hlavnímu predátorovi na Paramecium, mohou sloužit k fixaci živočicha na místě při krmení. U některých řasinek působí trichocysty jako útočné organely. Po vypuštění trichocyst se regenerují z kinetozomů.


(ix) Neuromotorika a přidružená vlákna:
Podle Lunda (1933) se na levé hřbetní stěně cytofaryngu přibližně na úrovni zadního okraje cytostomu nachází velmi malá, dvoubuněčná hmota, neuromotorium. Z neuromotoria vyzařují vlákna do endoplazmy.
Z nich čtyři nebo více obvykle procházejí téměř k hřbetní stěně těla, ale ostatní jsou kratší a jejich poloha není určitá. Všechny se označují jako endoplazmatické fibrily. Jejich funkce není známa, ale mohou koordinovat potravní pohyby ústních řasinek. Fibrily mohou také poskytovat mechanickou oporu, elasticitu, kontraktilitu, vodivost a metabolický vliv.

(x) Endoplazma:
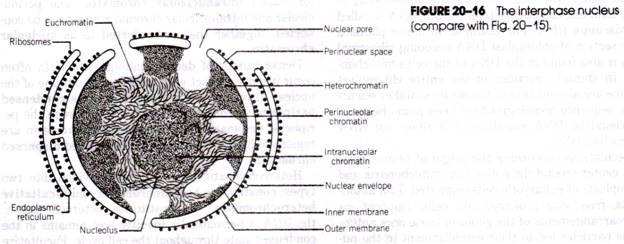
Endoplazma neboli dřeň je tekutější a objemnější část cytoplazmy, která obsahuje mnoho cytoplazmatických granulí a také další inkluze a struktury specializované povahy. Mezi cytoplazmatické inkluze patří mitochondrie, Golgiho aparát, vakuoly, krystaly, granula a chromidie atd. V endoplazmě se nacházejí také další struktury, a to jádra, kontraktilní vakuoly a potravní vakuoly.
Jádra:
V endoplazmě v blízkosti cytostomu jsou dvě jádra, tj. paramecium je heterokaryotické, velké elipsoidní a granulární makrojádro a další malé kompaktní mikrojádro.
Makronukleus je nápadné, elipsoidní nebo ledvinovité tělísko. Je kompaktního typu, obsahuje jemná vlákna a pevně zabalená diskrétní chromatinová zrnka různé velikosti a je uloženo v achromatické matrix. Má mnoho nukleolů a mnohem více chromatinového materiálu (DNA).
Jde o somatické nebo vegetativní jádro. Dělí se amitoticky a řídí vegetativní funkce (metabolické činnosti) živočicha. Podléhá však mitóze.
Mikrojádro je malé, kompaktní a kulovité. Obvykle se nachází v blízkosti makronuklea, často v konkavitě. Drobná chromatinová zrnka a vlákna jsou rovnoměrně rozmístěna v celé struktuře. Mikrojádro se dělí mitoticky a řídí reprodukci.
Počet mikrojader se liší podle druhu; u P. caudatum je jedno, u P. aurelia dvě a u P. multimicronucleatum mnoho. Mikrojádro obsahuje zřetelný nukleolus u P. aurelia, ale u P. caudatum se nevyskytuje. Moses (1949; 1950) uvádí, že makrojádro a mikrojádro mají shodné chemické složení.
(xi) Kontrakční vakuoly:
Existují dvě velké, tekutinou naplněné kontraktilní vakuoly, z nichž každá se nachází u jednoho konce těla blízko hřbetního povrchu. Jejich poloha je stálá (na rozdíl od améby), leží mezi ektoplazmatem a endoplazmatem, ale jsou to dočasné organely, které periodicky zanikají. U některých druhů se zdá, že mají výstelkovou membránu, a v takovém případě během systoly zcela nezmizí.
Ke každé kontraktilní vakuole je připojeno pět až dvanáct trubicovitých paprsčitých kanálků, z nichž každý se skládá z koncové části, dlouhé ampuly, která se při vyprázdnění zavírá, a krátkého vstřikovacího kanálku, který ústí do vakuoly.

Kanálky komunikují s velkou částí těla, odkud přijímají tekutiny a vlévají je do vakuoly, která se takto obnovuje a zvětšuje, když kontraktilní vakuola dosáhne maximální velikosti, náhle se stáhne (systola) a vypustí svůj obsah trvalým pórem v pelikulu, pak kanálky opět vytvoří kontraktilní vakuolu, kanálky zcela nezmizí, protože jsou to trvalé struktury.
Dvě kontraktilní vakuoly se vyprazdňují nepravidelně, zadní se stahuje rychleji, protože je blízko cytoplazmy a přichází do ní více vody. Hlavní funkce kanálků a kontraktilních vakuol je hydrostatická, odstraňují přebytek vody z protoplazmy, voda je částečně absorbována a částečně přijímána při krmení.
Dusíkaté odpadní látky mají amoniakální sloučeniny a některé uráty, které jsou vylučovány ze stahovacích vakuol spolu s CO2, ale nejsou žádné důkazy o tom, že by vylučovací látky byly vylučovány protoplazmou do kanálků nebo stahovacích vakuol.
Pod peristomem na jedné straně cytofaryngu je pevný stálý cytopyge nebo anus, kterým jsou vylučovány nestrávené zbytky potravy a uráty.
Elektronově mikroskopické studium kontraktilních vakuol odhalilo, že každý kontraktilní aparát se skládá z některých tubulů endoplazmatického retikula, nefridiálních tubulů, podávacích kanálků, akcesorních vakuol (radiálních kanálků) a hlavní kontraktilní vakuoly. Předpokládá se, že akcesorní vakuoly jsou ampulami podávacích kanálků (obr. 20.11).
(xii) Potravní vakuoly:
Jsou to zhruba kulovitá, nekontraktilní tělíska různé velikosti a počtu ležící v endoplazmě. Obsahují částice pozřené potravy, hlavně bakterie a malé množství tekutiny ohraničené tenkou definitivní membránou. Volkonsky (1934) navrhl pro tyto vakuoly název gastrioly. S potravními vakuolami jsou spojena trávicí granula.
Lokomoce Paramecium caudatum:
Paramecium caudatum vykonává lokomoci dvěma způsoby, a to metabolismem nebo konturami těla a pomocí řasinek.
(i) Metaboly neboli tělesné kontorze:
Tělo Paramecium caudatum má pružnost, může se protáhnout průchodem užším než jeho tělo, načež tělo nabývá normálního tvaru. Tato dočasná změna tvaru těla je metabolismus, u Paramecia ji vyvolává protoplazma.
(ii) Řasinkový pohyb:
Lokomoce vyvolaná řasinkami je hlavním způsobem. Řasinky mohou bít dopředu nebo dozadu, což umožňuje živočichovi plavat dopředu nebo dozadu.
Obvykle živočich plave dopředu, řasinky bijí dozadu, ale šikmo, řasinky ztuhnou a rychle se ohnou dozadu, až se téměř dotknou povrchu těla, tento tah se nazývá účinný tah; pak řasinky ochabnou a pomalu se vracejí do původní svislé polohy, to je , tzv. zotavovací tah.
Řasinky stejné příčné řady bijí společně a řasinky stejné podélné řady bijí jedna za druhou od předního k zadnímu konci.

Tento koordinovaný pohyb řasinek se nazývá metachronní rytmus, který je způsoben infraciliárním systémem; ten způsobuje plavání zvířete vpřed. Když však tělní řasinky bijí šikmo dozadu, pak zároveň delší řasinky ústní drážky bijí energičtěji, což způsobí, že se přední konec stočí doleva.
Působením řasinek tělní a ústní drážky se živočich otáčí kolem své dlouhé osy. Toto otáčení je vždy vlevo (s výjimkou P. calkinsi, který se otáčí po pravotočivé spirále).
Tato kombinace pohybu vpřed, otáčení a rotace způsobuje, že se živočich pohybuje vpřed po spirále proti směru hodinových ručiček. Tato dráha má přímou osu a stejný povrch těla zvířete zůstává směrem k ose spirálové dráhy. Při plavání vzad se však všechny druhy otáčejí doprava.
Ryt řasinek může být obrácený, takže se řasinky pohybují šikmo dopředu, čímž živočich plave dozadu. Činností řasinek se Paramecium pohybuje rychlostí 1500 mikronů nebo i více za sekundu.
Jennings tvrdí, že spirálovitost Paramecia je způsobena tím, že řasinky sice dopadají hlavně dozadu, ale šikmo doprava, čímž způsobují, že se živočich převrací doleva.
Také toto stáčení těla směrem k aborálnímu povrchu je do značné míry způsobeno větší silou účinného tahu ústních řasinek, které dopadají více přímo dozadu. Výsledek – otáčení paramecia kolem své dlouhé osy – tak umožňuje parameciu sledovat víceméně přímý směr při tvorbě velkých spirál.
Výživa Paramecium caudatum:
U Paramecium caudatum je výživa holozoická. Potrava se skládá především z bakterií a drobných prvoků. Paramecium na potravu nečeká, ale aktivně ji loví.
Uvádí se, že Paramecium caudatum vykazuje při výběru potravy selektivitu, ale zdá se, že pro to není žádný důvod, ačkoli pohlcuje pouze určité druhy bakterií; dostupné údaje naznačují, že jediné Paramecium pozře za 24 hodin 2 až 5 milionů jedinců Bacillus coli. Živí se také jednobuněčnými rostlinami, jako jsou řasy, rozsivky atd., a malými kousky živočichů a zeleniny.
Mechanismus výživy:
Když se Paramecium caudatum dostane do oblasti s hojnou potravou, odpočívá. Živí se pouze v klidu nebo při velmi pomalém plavání, při rychlém plavání se nikdy neživí. Tlukot řasinek ústní drážky způsobuje, že kuželovitý vír vody s potravou je vháněn do ústní drážky ze vzdálenosti před předním koncem (obr. 20.16).
Částice potravy se pak dostávají do předsíně, odkud jsou některé částice potravy odmítnuty a vyvrženy, ale jiné přecházejí do cytostomu.
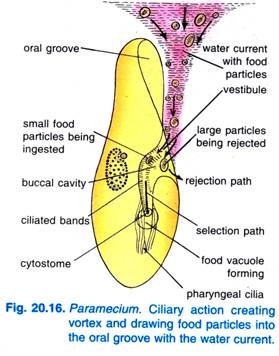
Na konci cytofaryngu se vytváří potravní vakuola, která se naplní částicemi potravy. Kvadrulus a peniculi řídí průchod potravy do potravní vakuoly, která se vytváří laterálně. Když potravní vakuola dosáhne určité velikosti, sevřou ji postbulbární vlákna, která ji přimáčknou a vydají se na cestu.
Vakuola obsahuje kromě potravy také trochu vody. Rotační proudové pohyby endoplazmy zvané cyklóza nesou potravní vakuoly po určité dráze, která funkčně odpovídá trávicímu traktu.
Dráha začíná od konce cytoplazmy, pak na zadní stranu, pak dopředu, aby cirkulovala s endoplazmatem, pak na hřbetní povrch, pak směrem k přednímu konci, pak dolů k cytoplazmě. Na počátku své cesty se velikost potravní vakuoly zmenšuje, pak se opět zvětšuje.

Trávení a vylučování Paramecium Caudatum:
Během cyklózy dochází k trávení pomocí enzymů vylučovaných protoplazmou do vakuol. Při trávení se bílkoviny mění na aminokyseliny, sacharidy na rozpustné cukry a glykogen a pravděpodobně se tráví i tuky.
Obsah potravních vakuol je nejprve kyselý (pH asi 4) a poté se stává zásaditým, hlavní trávení probíhá během zásadité fáze. Nestrávená hmota je určitou silou vyvržena přes cytopyge.
Cyklózu lze prokázat experimentálně; pokud se Parameciu podá mléko obarvené konžskou červení, tukové kuličky mléka v potravních vakuolách nejprve zčervenají v důsledku kyselé reakce enzymů, poté se změní z odstínů fialové na modrou v důsledku alkalické reakce, vakuoly vykazují průběh cyklózy.
Dýchání a vylučování Paramecium caudatum:
Výměna plynů (kyslíku a oxidu uhličitého) probíhá přes polopropustnou pelikulu stejně jako u jiných sladkovodních prvoků procesem difuze. Paramecium Caudatum získává kyslík z okolní vody. Oxid uhličitý a organické odpadní látky, jako je amoniak, vznikající při metabolismu, jsou pravděpodobně vylučovány difuzí ven do vody v opačném směru.

6. Jak probíhá dýchání, vylučování a osmoregulace? Osmoregulace u Paramecium Caudatum:
Paramecium Caudatum má dvě kontraktilní vakuoly, jednu přední a jednu zadní. Funkcí kontraktilních vakuol je osmoregulace, tj. regulace obsahu vody v těle, a mohou sloužit také k vylučování dusíkatých odpadních látek, jako je močovina a amoniak.
Přebytek vody (v důsledku nepřetržité endosmózy) v cytoplazmě se vylučuje do kanálků endoplazmatického retikula a přechází do nefridiových kanálků → podávacích kanálků → a shromažďuje se v ampulích řady 6 až 11 paprsčitých kanálků, které se sbíhají směrem ke každé vakuole a vypouštějí ji do ní. Kanály jsou nejvíce patrné, když se tvoří vakuola.
Když každá vakuola nabobtná (diastola) do určité velikosti, smrští se (systola) a vypouští se ven pravděpodobně pórem. Kontrakční vakuoly se stahují střídavě, v intervalech 10 – 20 sekund.
Zadní kontraktilní vakuola pracuje rychleji než přední vakuola, protože cytoplazma přijímá velké množství vody do zadní oblasti. Kontraktilní vakuoly udržují optimální koncentraci vody v cytoplazmě těla tím, že se zbavují jejího přebytku.

Chování Paramecium caudatum:
Odezvy Paramecium caudatum na různé druhy podnětů se poznají studiem jeho reakcí a seskupení nebo rozptýlení jedinců v kultuře. Reakce je pozitivní, pokud se živočich pohybuje směrem k podnětu, a negativní, pokud se vzdaluje. Na nepříznivý podnět zvíře reaguje vyhýbavě, dokud neuteče.
Při vyhýbavé reakci se ciliární rytmus obrátí, zvíře se posune o krátkou vzdálenost dozadu a pak se otáčí po kuželovité dráze tak, že přední konec vychýlí aborálně, zatímco se otáčí na zadním konci. Všechny úpravy se provádějí metodou pokusu a omylu. Pokusy ukázaly, že přední konec zvířete je citlivější než ostatní části.
Reakce paramecií na různé podněty lze rozdělit do následujících skupin:
(i) Reakce na kontakt (Thigmotaxis):
Reakce na kontakt jsou u Paramecia různé. Pokud se předního konce lehce dotkneme jemným hrotem, dochází k silné vyhýbavé reakci. Při plavání Paramecium narazí na nějaký předmět ve vodě, ale pokud se dotkne jiného místa, nemusí dojít k žádné reakci. Pomalu se pohybující jedinec často reaguje, pozitivně na kontakt s nějakým předmětem tím, že na něm spočine.
(ii) Reakce na chemické látky (chemotaxe):
Všeobecně Paramecia reagují na chemické podněty tak, že se vyhýbají reakci. Pokud se do populace Paramecií na mikroslidu vnese kapka slabého roztoku soli (0,5 %), živočichové reagují vyhýbavou reakcí a žádný z nich do kapky nevstoupí. Na kyseliny však reagují pozitivně, i když je koncentrace dostatečně silná, aby je usmrtila.
(iii) Reakce na teplotu (termotaxe):
Paramecium vyhledává optimální teplotu 24 až 28 °C. Pokud dojde ke změně teploty výrazně nad nebo pod optimální rozmezí, Paramecia vykazují vyhýbavou reakci. Větší teplo stimuluje rychlý pohyb a vyhýbání se reakcím, dokud zvířata neutečou nebo nejsou usmrcena.
(iv) Reakce na světlo (fototaxe):
S výjimkou zeleného Paramecium bursaria, které je pozitivně fototaktické, jsou ostatní druhy k běžnému světlu lhostejné. Při náhlém a prudkém zvýšení intenzity světla však zpravidla následuje negativní reakce. Paramecia vykazují okamžitou negativní reakci na ultrafialové záření.
(v) Reakce na elektrický proud (galvanotaxe):
Paramecia reagují na elektrické podněty. Když se v mělké misce s Paramecia umístí dvě elektrody proti sobě a pustí se do nich konstantní proud, všechny organismy plavou stejným směrem ke katodové neboli záporné elektrodě, kde se koncentrují ve velkém množství.
Pokud se směr elektrického proudu změní, zatímco Paramecia plavou ke katodě, organismy obrátí směr a plavou k nové katodě.

(vi) Reakce na vodní proud (reotaxe):
Paramecia vykazují pozitivní reotaxi. V mírném vodním proudu se paramecie většinou pohybují s proudem svými předními konci proti proudu.
(vii) Reakce na gravitaci (geotaxe):
Paramecia obecně vykazují negativní reakci na gravitaci, jak je vidět v kultuře, kde se mnoho jedinců shromažďuje těsně pod povrchovou vrstvou s předními konci nahoře. Pokud jsou paramecie zavedeny do obrácené vodou naplněné zkumavky ve tvaru písmene U, která je na obou koncích zazátkována, okamžitě se přesunou nahoru do vodorovné části zkumavky.
Reprodukce u Paramecium Caudatam:
Paramecium Caudatum se rozmnožuje nepohlavně příčným binárním dělením a také prochází několika typy jaderné reorganizace, jako je konjugace, endomixis, autogamie, cytogamie a hemixis atd.
(i) Příčné binární dělení:
Příčné binární dělení je nejběžnějším typem nepohlavního rozmnožování u Paramecium. Je to výrazně unikátní asexuální proces, při kterém se jeden plně dospělý jedinec rozdělí na dva dceřiné jedince, aniž by zanechal rodičovské tělo.
Rovina dělení prochází středem buňky a probíhá v rovině kolmé na dlouhou osu těla. Dělení buněčného těla jako celku vždy předchází dělení jader; zdá se totiž, že rozmnožování je iniciováno jadernou aktivitou a dělením.

Paramecium caudatum se za příznivých podmínek rozmnožuje příčným binárním dělením. Při binárním dělení se mikrojádro rozdělí mitózou na dvě dceřiná mikrojádra, která se přesunou na opačné konce buňky. Makronukleus se prodlužuje a dělí se příčně amitózou.
Další cytoplazma se oddělí a objeví se dvě nové kontraktilní vakuoly, jedna u předního konce a druhá u zadního konce. Mezitím se poblíž středu těla objeví konstrikční brázda, která se prohlubuje, až je cytoplazma zcela rozdělena.
Vzniklá dvě „dceřiná“ paramecia jsou stejně velká a každé obsahuje sadu buněčných organel. Ze dvou vzniklých dceřiných paramécií se přední nazývá proter a zadní opisthe. Než dojde k dalšímu dělení, dorostou do plné velikosti.
Proces binárního dělení trvá přibližně dvě hodiny a může k němu dojít jednou až čtyřikrát za den, přičemž vzniká 2 až 16 jedinců. Za rok vznikne asi 600 generací.
Rychlost množení závisí na vnějších podmínkách potravy, teploty, stáří kultury a hustotě populace; také na vnitřních faktorech dědičnosti a fyziologie. Přirozeně, kdyby všichni potomci jednoho jedince přežili a rozmnožili se, počet vyprodukovaných paramécií by se brzy rovnal objemu země.
Termín klon se používá pro označení všech jedinců, kteří vznikli z jednoho jedince štěpením. Všichni členové klonu jsou si dědičně podobní.
(ii) Konjugace:
Obvykle se Paramecium caudatum množí binárním dělením po dlouhou dobu, ale v určitých intervalech může být tento proces přerušen spojením dvou jedinců podél jejich ústních ploch za účelem pohlavního procesu konjugace.
Konjugace je definována jako dočasné spojení dvou jedinců, kteří si vzájemně vyměňují mikrojaderný materiál. Je to jedinečný typ pohlavního procesu, při kterém se dva organismy oddělí brzy po výměně jaderného materiálu.
Sonneborn (1947) na základě pářícího chování Paramecium Caudatum uvedl, že každý druh Paramecium existuje v několika varietách neboli syngenech. Dále v rámci každého syngenu existuje několik typů páření, obvykle dva.
Typy páření zůstávají morfologicky shodné, ale vykazují fyziologické rozdíly. U P. aurelia existuje 14 syngenů a 28 typů páření, zatímco u P. caudatum existuje 16 syngenů a 32 typů páření. Bylo zjištěno, že paramecia obvykle nekonjugují ani s příslušníky vlastního pářícího typu, ani s jinými varietami, ale pouze s druhým pářícím typem vlastní variety.

Faktory vyvolávající konjugaci:
Faktory vyvolávající konjugaci se u jednotlivých druhů liší, ale některé z nich jsou uvedeny níže:
1. Ke konjugaci dochází obvykle za nepříznivých životních podmínek; uvádí se, že hladovění nebo nedostatek potravy a zvláštní bakteriální strava nebo určité chemické látky vyvolávají proces konjugace u některých druhů Paramecium.
2. Ke konjugaci dochází asi po 300 asexuálních generacích binárního dělení, nebo se střídá s binárním dělením v dlouhých intervalech, aby se omladil odumírající klon, tj. dochází k ní u jedinců, kteří museli projít žádoucím počtem asexuálních generací, což je prý období nezralosti, a pak pohlavně dozrají ke konjugaci.
3. Ke konjugaci dochází, když dojde ke změně fyziologického stavu paramécií, pak k ní dochází mezi takovými jedinci, kteří jsou poněkud menší velikosti (210 mikrometrů dlouzí) a jsou ve stádiu, které lze považovat za období nezdravého stáří; paramécie tohoto stavu uhynou, pokud jim není umožněno konjugovat.
4. Uvádí se, že náhlá tma ve světelných podmínkách a nízké teploty vyvolávají u některých druhů proces konjugace.
5. Konjugace neprobíhá v noci nebo za tmy; začíná brzy ráno a pokračuje až do odpoledne.
6. Říká se, že bílkovinná látka v řasinkách jedinců pářícího se typu vyvolává konjugaci.
Proces konjugace:
Proces konjugace se u různých druhů Paramecium liší, ale níže uvedený popis se týká procesu konjugace u P. caudatum (obr. 20.21).
Při konjugaci se dvě Paramecium caudatum (označované jako prekonjuganty) opačných pářících se typů téhož druhu přiblíží svými ventrálními povrchy a spojí se ústními rýhami; jejich řasinky produkují na povrchu těla látku, která způsobuje adhezi obou konjugujících paramécií.
Přestávají se živit a jejich ústní rýhový aparát zaniká. Pelikuly a ektoplazma se v místě styku obou rozpadají a mezi oběma živočichy vzniká protoplazmatický most. Nyní se tito jedinci nazývají konjuganti.
V tomto stavu konjugující pár aktivně plave a současně u každého konjuganta dochází k řadě jaderných změn, jak je popsáno níže:
Makronukleus se začíná rozpadat, stává se volným ve struktuře a vytváří složité stočené klubko, během druhé poloviny období konjugace nakonec zmizí, přičemž je pohlcen cytoplazmou. Mikrojádro každého konjugátu se dělí dvakrát, přičemž jedno z nich je redukční dělení.
V každém konjugátu tak vznikají čtyři haploidní dceřiná mikrojádra. Tři z těchto čtyř mikrojader v každém z nich degenerují, takže zůstane pouze jedno.
Zbylé mikrojádro každého konjugantu se mitoticky rozdělí na dvě nestejná pronuklea neboli herní jádra a vytvoří větší stacionární samičí pronukleus a menší aktivní migrující samčí pronukleus.
Migrující projádro jednoho konjugantu přechází přes protoplazmatický most a splývá se stacionárním projádrem druhého konjugantu, čímž vzniká synkaryon neboli konjugační jádro, v němž se obnovuje diploidní počet chromozomů a došlo k výměně dědičného materiálu.
Proces byl srovnáván s oplozením u vyšších živočichů, ale nejedná se o oplození, protože se nejedná o gamety. Konjuganti se nyní (asi po 12-48 hodinách) oddělují a nazývají se exkonjuganti. Synkaryon každého ex-konjugantu se třikrát rozdělí a vytvoří osm mikrojader v každém ex-konjugantu.
Čtyři z osmi mikrojader se zvětší a stanou se makrojádry a tři z ostatních čtyř mikrojader zaniknou.
Zbylé mikrojádro se rozdělí a současně se exkonjugant rozdělí binárním dělením na dvě buňky, z nichž každá má dvě makrojádra a jedno mikrojádro. Buňky a jejich mikrojádra se podruhé rozdělí a z každého exkonjugantu vzniknou čtyři paramecia, takže každé má jedno makrojádro a jedno mikrojádro.
Nové makrojádro, stejně jako mikrojádro, bylo vytvořeno z nového materiálu. Tato nová jádra pravděpodobně obsahují nový a odlišný potenciál, který se projevuje u zdravých jedinců.
Význam konjugace:
Klon odumře, pokud nedojde k jaderné reorganizaci, ale klon může být omlazen, aby získal zpět svou dřívější vitalitu pomocí jaderné reorganizace, tato jaderná reorganizace je vyvolána konjugací, konjugace je tedy nezbytná pro pokračování binárního dělení.
Význam konjugace byl shrnut níže:
1. Konjugace slouží jako proces omlazení a reorganizace, kterým se obnovuje vitalita rasy. Pokud ke konjugaci dlouhodobě nedochází, paramecia slábnou a odumírají. (Woodruffovo tvrzení o zachování zdravých paramécií po 22 000 generací bez konjugace vyvrátil Sonneborn, protože ukázal, že všechny Woodruffovy paramécie patřily ke stejnému párovacímu typu).
2. U konjugantů se nerozlišuje pohlaví, ačkoli konjugují pouze paramecia dvou různých pářících se typů téže variety.
3. Pohlaví se nerozlišuje, přesto se aktivní migrující projádro považuje za samčí a stacionární projádro za samičí.
4. Konjugace je pouze dočasné spojení, nedochází ke splynutí cytoplazmy a nevzniká zygota, ale jádro každého exkonjuganta obsahuje dědičný materiál dvou konjugujících jedinců.
5. Konjugací dochází k nahrazení makrojádra materiálem ze synkaryonu, to je událost zásadního významu. Při binárním dělení byly chromozomy makronukleu náhodně rozděleny do dceřiných buněk, pokračující binární dělení způsobilo, že klon byl slabý s některými strukturními abnormalitami.
Konjugací dochází k vytvoření správného počtu chromozomů v makronukleu, takže se obnovuje rasa v síle. Úkolem mikrojádra je obnovit vyvážený komplex chromozomů a genů.
Aberantní chování při rozmnožování u Paramecium Caudatum:
Paramecium Caudatum vykazuje určité odchylky v jaderném chování během dělení a konjugace, těmito odchylkami jsou endomixis, autogamie, cytogamie a hemixis. Při prvních třech procesech dochází ke genetické rekombinaci a z mikrojádra vzniká nové makrojádro.
(i) Endomixe:
Woodruff a Erdmann (1914) jako první popsali nový proces jaderné reorganizace, endomixis (gr., endon = uvnitř; mixis = míšení) u Paramecium aurelia, bimikronukleárního druhu (obr. 20.22). Tento proces byl popsán jako periodicky probíhající, při němž vzniká nový makrojaderný aparát bez tvorby synkaryonů. K endomixi dochází v rámci jednoho jedince.
Podle Woodruffa a Erdmanna makronukleus degeneruje a mikrojádra se dělí dvakrát za vzniku osmi mikrojader. Šest mikrojader degeneruje a dvě zůstávají. Když zůstanou pouze dvě mikrojádra, živočich se rozdělí štěpením na dvě buňky, každou s jedním mikrojádrem.
Mikrojádro každé dceřiné buňky se dělí dvakrát a vytváří čtyři mikrojádra. Dvě mikrojádra se zvětší a vytvoří makrojádra. Živočich a jeho mikrojádra se rozdělí tak, že vzniknou dva dceřinní jedinci, z nichž každý má jedno makrojádro a dvě mikrojádra.
Endomixe se vyskytuje u té odrůdy P. aurelia, která nekonjuguje, tudíž účinek endomixe může být stejný jako účinek konjugace, protože oba procesy vedou k nahrazení makrojádra materiálem z mikrojádra a oba procesy omlazují vitalitu rasy.
Ale oba procesy se liší, protože při endomixi nedochází ke splynutí pronukleů; endomixi lze přirovnat k partenogenezi.
Někteří pracovníci však oprávněně tvrdí, že endomixe není platným procesem a byla popsána v důsledku chybného pozorování. Se vší pravděpodobností k endomixi nedochází a může se jednat pouze o specializovaný případ autogamie.

Později Erdmamm a Woodruff (1916) zaznamenali endomixi u Paramecium caudatum. Diller však nevěří v platnost tohoto procesu a domnívá se, že Erdmann a Woodruff jednoduše spojili stadia hemixie a autogamie do jednoho schématu, endomixie.
(ii) Autogamie:
Diller (1934, 1936) a Sonneborn (1950) popsali proces samooplození neboli autogamie probíhající u jediného jedince u Paramecium aurelia (obr. 20.23). Uvedl, že při autogamii vznikají při třech mikrojaderných (pregamických) děleních, zahrnujících zrání, herní jádra (pronuklea).
Během autogamie u P. aurelia se dvě mikrojádra dělí dvakrát (jednou meioticky) a vzniká osm mikrojader, z nichž šest degeneruje. Mezitím se makronukleus rozroste ve skeletovitou hmotu, která se později rozpadne na kousky, které se vstřebají do cytoplazmy. Dvě z osmi mikrojader se jako pronuklea dostanou do protoplazmatického kužele vybouleného poblíž buněčného ústí.
Dvě pronuklea splynou a vytvoří synkaryon. Synkaryon se dvakrát rozdělí a vytvoří čtyři mikrojádra. Ze dvou mikrojader se stanou makrojádra. Paramecium a jeho mikrojádra se rozdělí za vzniku dvou dceřiných jedinců, z nichž každý má jedno makrojádro a dvě mikrojádra. Tento proces je dokončen přibližně za dva dny.
Autogamie přináší omlazení rodu.
Podobá se konjugaci v tom, že nové makrojádro vzniká z materiálu mikrojádra, v novém makrojádru se obnovuje správný počet chromozomů; a také v tom, že dochází ke splynutí dvou pronukleů. Autogamie se však od konjugace liší tím, že autogamie se účastní pouze jeden jedinec, který poskytuje obě pronuklea, jedná se o druh samooplození.
(iii) Hemixis:
Diller (1936) popsal hemixii u Paramecium aurelia (obr. 20.24). Hemixis je především proces makrojaderné fragmentace a dělení bez neobvyklé mikrojaderné aktivity. Diller rozdělil hemixi do čtyř typů, a to A, B, C a D, jak ukazuje obr. 20.24 u P. aurelia, ale se všemi typy se setkal i v masových kulturách P. caudatum a P. multimicronucleatum.
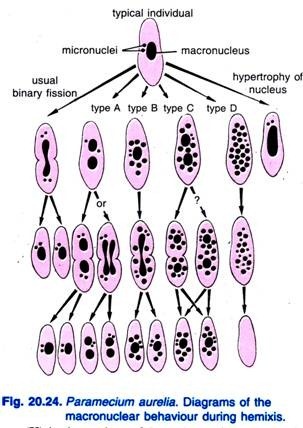
Typ A je nejjednodušší forma hemixe charakterizovaná rozdělením makrojádra na dvě nebo více částí. Toto dělení není synchronizováno s dělením mikrojader.
Typ B je charakterizován vytlačením jedné až 20 nebo více chromatinových kuliček z makronuklea do cytoplazmy.
Typ C je charakterizován současným rozdělením makrojádra na dvě nebo více hlavních částí a vytlačováním makronukleárních kuliček do cytoplazmy.
Typ D je považován za představitele patologických stavů, při nichž makronukleus prochází úplnou fragmentací na chromatinové kuličky, které nakonec z buňky zmizí. Mikrojádra obvykle zanikají před rozpuštěním makrojádra.
(iv) Cytogamie:
Wichterman (1939) popsal další sexuální proces u Paramecium caudatum, který nazval cytogamie. Při cytogamii nedochází k výměně jader. Při tomto procesu se dva jedinci spojí svými ventrálními povrchy, ale pelikuly obou jedinců se nerozpadají.
Mikrojádro každého jedince se třikrát dělí a vytváří osm mikrojader, z nichž šest se rozpadá v každém jedinci. Dvě zbývající mikrojádra splynou a vytvoří v každé buňce synkaryon. Jedinci se nyní oddělí.
Cytogamie se liší od autogamie tím, že jsou dva jedinci ve vzájemném kontaktu, ale podobá se autogamii a konjugaci splynutím dvou pronukleí. Cytogamie se od konjugace liší tím, že nedochází k výměně jader mezi dvěma živočichy, kteří se spojují.
Některé cytoplazmatické částice popsané u Paramecium caudatum:
(i) částice kappa:
V roce 1938 T. M. Sonneborn uvedl, že některé rasy (známé jako zabijáci nebo zabijácký kmen) Paramecium produkují jedovatou látku zvanou paramecin, která je smrtelná pro jiné jedince zvané citlivci. Paramecin je rozpustný ve vodě, difúzní a jeho produkce závisí na některých částicích nacházejících se v cytoplazmě Paramecia (kmene killer).
Tyto částice se nazývají částice kappa. Částice kappa mají DNA a RNA. Zabijácké Paramecium může obsahovat stovky částic kappa. Podrobné studium těchto částic odhalilo, že pro existenci částic kappa, jejich množení a produkci paramecinu je nezbytný dominantní gen (K) v jádře Paramecia.
(ii) mµ, částice:
R.W. Siegel (1952) popsal další typ zabijáckých částic v cytoplazmě některých Paramecií. Paramecium s částicemi mµ se nazývá mate killer, protože když konjuguje s parameciem bez částic mµ, které se nazývá mate sensitive, pak toto paramecium zabíjí. Částice mp jsou také složeny z DNA, RNA atd.
Tyto částice existují pouze u těch paramécií, jejichž mikrojádro obsahuje alespoň jeden dominantní gen jednoho ze dvou párů nespojitých chromozomálních genů (M1 a M2).
(iii) Částice pí:
Tyto částice mají být mutantní formou částic kappa, ale neprodukují žádný typ jedovaté látky.
(iv) Částice lambda:
Tyto částice jsou uváděny u zabijáckých paramécií a údajně produkují nějakou látku odpovědnou za způsobení lýzy nebo rozpadu citlivých paramécií, tj. těch, které ji nemají.