Gameten und Befruchtung Übersicht
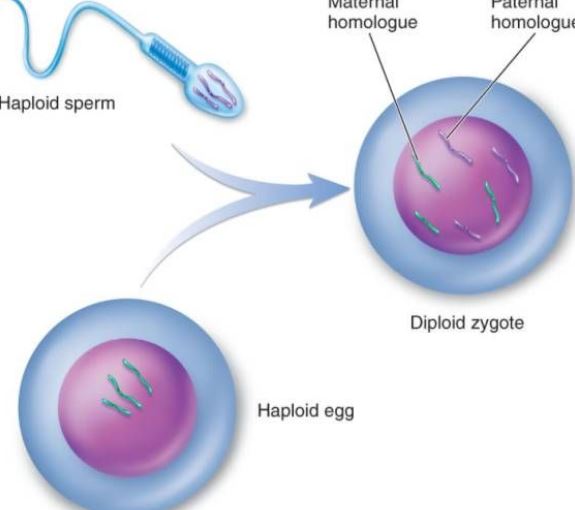
Die sexuelle Fortpflanzung bezieht sich auf die von den Eltern produzierten geschlechtlichen Keimzellen (Gameten), die durch die Kombination von zweigeschlechtlichen Keimzellen (wie Spermien und Eizellen) zur befruchteten Eizelle werden, und dann entwickelt sich die befruchtete Eizelle zu einem neuen Individuum, was als sexuelle Fortpflanzung bezeichnet wird. Eine umfangreiche Variation der Genkombinationen bei der sexuellen Fortpflanzung kann die Fähigkeit der Nachkommen erhöhen, sich der natürlichen Selektion anzupassen. Zufällig kombinierte Gene in den Nachkommen der sexuellen Fortpflanzung können für die Art vorteilhaft sein oder auch nicht, erhöhen aber zumindest die Chancen einiger weniger Individuen, in einer unvorhersehbaren und sich ständig verändernden Umwelt zu überleben, und kommen damit der Art zugute. Die sexuelle Fortpflanzung kann auch die Verbreitung günstiger Mutationen in der Population fördern. Wenn zwei Individuen einer Art an verschiedenen Stellen günstige Mutationen aufweisen, konkurrieren die beiden Mutanten in der Population der ungeschlechtlichen Fortpflanzung so lange miteinander, bis sie eliminiert werden, denn es ist unmöglich, beide günstigen Mutationen gleichzeitig zu erhalten. In Populationen, die sich sexuell fortpflanzen, können diese beiden günstigen Mutationen jedoch durch Paarung und Rekombination gleichzeitig in das Genom desselben Individuums gelangen und sich gleichzeitig in der Population verbreiten. Aus den oben genannten Gründen beschleunigt die sexuelle Fortpflanzung den Evolutionsprozess. In den mehr als 3 Milliarden Jahren der biologischen Evolution auf der Erde blieben die ersten zwei Milliarden Jahre des Lebens im Stadium der ungeschlechtlichen Fortpflanzung, und die Evolution verlief langsam, und seit den letzten 1 Milliarde Jahren hat sich die Evolutionsrate erheblich beschleunigt. Neben Veränderungen in der Umwelt der Erde (wie dem Aufkommen der sauerstoffhaltigen Atmosphäre usw.) ist auch das Auftreten und die Entwicklung der sexuellen Fortpflanzung eine wichtige Ursache.
Gametes
Gametes sind reife Zellen, die vom Fortpflanzungssystem produziert werden, wenn der Organismus sich sexuell fortpflanzt und die als Keimzellen bezeichnet werden. Gameten werden in männliche und weibliche Gameten unterteilt. Die weiblichen Geschlechtszellen von Tieren und Pflanzen werden in der Regel als Eizellen bezeichnet, die männlichen Geschlechtszellen als Spermien. Spermien sind recht klein, können sich aber bewegen und skorpionförmig in die Eizelle eindringen. Die Eizelle ist recht groß und nicht schwimmfähig. Beim Seeigel beispielsweise ist das Volumen der Eizelle 10.000 Mal größer als das der Samenzellen. Obwohl die männlichen und weiblichen Geschlechtszellen ein unterschiedliches Volumen haben, ist die Kern-DNA, die sie für die Nachkommenschaft bereitstellen, gleich, d. h. jede liefert einen Satz von Genomen. Da die Eizellen jedoch sehr groß sind, werden die zytoplasmatische Struktur und die zytoplasmatische DNA der Tochterzellen hauptsächlich von den Eizellen bereitgestellt. Keimzellen spielen eine wichtige Rolle bei biologischen Berechnungen. Anhand von Genkarten lassen sich der Genfluss und das Muster des Genotyps der Nachkommen deutlich erkennen. Die Anzahl der Gameten in der ersten Generation von Hybriden und die relativen Merkmale in Hybriden sind eine 2n-te Potenz (n ist der Logarithmus der relativen Merkmale). Wenn ein Lebewesen eine Gamete bildet, werden die gepaarten genetischen Faktoren voneinander getrennt und gelangen in verschiedene Gameten. Die Gameten enthalten von jedem Paar genetischer Faktoren nur eines. Bei der Befruchtung ist die Kombination von männlichen und weiblichen Gameten zufällig. Aufgrund der Gattung der durch Meiose gebildeten Gameten, der Vielfalt der Chromosomenzusammensetzung, der Unterschiede im genetischen Material der verschiedenen Gameten und der Zufälligkeit der Kombination von Eizellen und Spermien bei der Befruchtung müssen die Nachkommen der gleichen Eltern unterschiedlich sein.
Befruchtung
Die Befruchtung ist der Prozess, bei dem Eizellen und Spermien zu einer Zygote verschmelzen. Sie ist ein grundlegendes Merkmal der sexuellen Fortpflanzung und ist in der Tier- und Pflanzenwelt allgegenwärtig, wird aber oft als die am häufigsten auf das Tier bezogene bezeichnet. Der Befruchtungsprozess bei Tieren umfasst auf zellulärer Ebene drei Hauptphasen: Aktivierung der Eizelle, Regulierung und amphotere Vorkernfusion. Die Aktivierung kann als Ausgangspunkt der Ontogenese angesehen werden, die sich vor allem in Veränderungen der Durchlässigkeit der Eimembran, in der Bildung von Rindenkörnchen, in der Bildung der befruchteten Membran usw. äußert. Die Anpassung erfolgt nach der Aktivierung, die der erste Schritt zur Gewährleistung der normalen Teilung der befruchteten Eizellen ist. Die Veränderung des prokaryotischen Zellkerns gewährleistet die Vererbung der Eltern und stellt die Diploidie wieder her. Durch die Befruchtung wird nicht nur die Replikation der DNA eingeleitet, sondern auch die genetische Information wie mRNA und rRNA in der Eizelle aktiviert, um das für die Entwicklung des Embryos erforderliche Protein zu synthetisieren. Nach 6 bis 7 Tagen nach der Befruchtung verschwindet die Zona pellucida der Blastozyste und wird allmählich von der Gebärmutterschleimhaut bedeckt, was als Einnistung der befruchteten Eizelle bezeichnet wird. Die Spermien eines Tieres haben keine offensichtliche Chemotaxis wie die Spermien niederer Pflanzen, z. B. der Moose, sondern erreichen die Nähe der Eizelle durch aktive Bewegung oder durch die Zilienbewegung der Genitalepithelzellen. Kondensation der Spermien: Es ist bekannt, dass bei der Passage vieler Säugetierspermien durch den weiblichen Fortpflanzungstrakt oder durch den Kumulus das Fremdprotein, das die Spermien einkapselt, entfernt wird und sich die physikalischen und biologischen Eigenschaften der Plasmamembran der Spermien ändern, so dass die Spermien Energie gewinnen und am Befruchtungsprozess teilnehmen können. Wenn das kapazitierte Säugetierspermium mit der Eimembran oder Zona pellucida der Eizelle in Kontakt kommt, bindet es spezifisch an ein Glykoprotein auf der Eimembran, das das Spermium zur Energiegewinnung anregt. Die Akrosomreaktion hilft dem Spermium, die Eimembran weiter zu durchdringen. Auf dem Seeigel-Ei ist die Akrosom-Reaktion der Spermien eine bestimmte Polysaccharid-Substanz in der Perimembranschicht. Die meisten Eier haben Eimembranen an der Peripherie, und die Dicke der verschiedenen Eimembranen ist unterschiedlich. Der Hauptbestandteil ist Mucin oder Mucopolysaccharid; nur wenige sind nackte Eier, wie die Eier eines Coelenteraten. Akrosom-Reaktion: Wenn das Spermium die Eimembran durchdringt, kommt es zu einem Bindungsprozess. Das Spermium haftet nur lose an der Eihülle, ist frei von äußeren Temperatureinflüssen und hat keine spezifische Eigenschaft. Während der Adhäsionsphase wird das Protoplastenprotein auf der Akrosomenmembran in das Akrosomenprotein umgewandelt, und das Akrosomenprotein beschleunigt die Spermien, die Eimembran zu durchqueren; es ist eine starke Verbindung, die durch niedrige Temperaturen gestört werden kann und spezifisch ist. Auf der Plasmamembran der Seeigelspermien wurde ein Protein isoliert, das spezifisch an das Glykoprotein der Eizelle bindet, ein so genanntes Bindungsprotein mit einem Molekulargewicht von etwa 30.000. Die sekundären Eizellen, die sich während des Eisprungs in der Mitte der zweiten Reifeteilung befinden, dringen zusammen mit der sie umgebenden Zona pellucida und der Strahlenkrone durch die Bauchhöhle rasch in die Eileiterampulle ein, was auf das Schwingen der Flimmerhärchen der Eileiterepithelzellen und die Kontraktion der Muskelschicht zurückzuführen ist. Trifft die Eizelle nicht auf das Spermium, beginnt sie in der Regel innerhalb von 12-24 Stunden zu degenerieren. Sobald das Spermium mit der Eizelle in Kontakt gekommen ist, durchläuft die Eizelle selbst eine Reihe von Aktivierungsänderungen. Bei Eizellen von Säugetieren ist dies durch die Rindenreaktion, die Ovarialmembranreaktion und die Zona-Pellucida-Reaktion gekennzeichnet, die die Mehrfachbefruchtung blockieren und die weitere Entwicklung der Eizellen stimulieren können. Die kortikale Reaktion tritt zum Zeitpunkt der Verschmelzung der Samenzellen auf. Ab dem Zeitpunkt der Verschmelzung reißen die Rindenpartikel auf, und der Inhalt tritt aus, wodurch sich die Rinde der gesamten Eizelle ausbreitet. Die Ovarialmembranreaktion ist ein Rekombinationsprozess der Eizelle und der Granulathülle der Rinde. Die Zona-Pellucida-Reaktion ist ein Prozess, bei dem der kortikale Efflux und die Zona Pellucida von einer befruchteten Membran abgetrennt werden, die Eimembran von der Plasmamembran getrennt wird, der Spermienrezeptor in der Zona Pellucida verschwindet und die Zona Pellucida verhärtet. Nur Spermien, die eine Akrosomreaktion zeigen, können mit der Eizelle verschmelzen. Unter der Einwirkung von Akrosin passiert das Spermium die radiale Krone und interagiert mit dem Spermienrezeptor-Glykoproteinmolekül ZP3 auf der Zona pellucida, wodurch das Spermium das akrosomale Enzym freisetzen und durch die Zona pellucida in die Perivitelline eintreten kann. Zu Beginn der Befruchtung ist die Hülle des äquatorialen Teils des menschlichen Spermienkopfes in Kontakt mit der Eizellmembran, und dann dringen der Kern und das Zytoplasma des Spermiums in die Eizelle ein. Nachdem das Spermium in die Eizelle eingedrungen ist, geben die Rindenpartikel im flachen Zytoplasma der Eizelle sofort ihren Inhalt an die Peripherie der Membran ab. Während die Membran der Rindenkörnchen mit den Eizellen verschmilzt, erhöht sich gleichzeitig die negative Oberflächenladung auf der Zelloberfläche, wodurch die Verschmelzung der Plasmamembran der Spermien mit der Eimembran unterdrückt wird, was als Rindenreaktion bezeichnet wird. Die Struktur der Zona pellucida verändert sich, was als Zona-pellucida-Reaktion bezeichnet wird. Zu diesem Zeitpunkt ist die Fähigkeit der Zona pellucida, Spermien zu binden, reduziert, wodurch das Auftreten von Polyspermie verhindert und die biologischen Merkmale der menschlichen Monospermie gewährleistet werden. Neben der Zerstörung oder Inaktivierung des an die Spermien in der Zona pellucida gebundenen Rezeptors kann der Inhalt der Rindengranula auch die Vernetzung zwischen den Peptidketten in der Zona pellucida erhöhen, die Empfindlichkeit gegenüber der Akrosomenprotease schwächen und das Eindringen der Spermien verhindern. Obwohl mehrere Spermien die Zona pellucida passieren, dringt nur ein Spermium in die Eizelle ein, um sie zu befruchten. In einer anormalen Situation sind zwei Spermien an der Befruchtung beteiligt, nämlich die Befruchtung durch zwei Spermien. Zwei Spermien dringen gleichzeitig in die Eizelle ein und bilden Embryonen aus triploiden Zellen, die alle abgetrieben werden oder kurz nach der Geburt sterben. Nachdem das Spermium in die Eizelle eingedrungen ist, vollzieht die Eizelle rasch die zweite reife Teilung. Zu diesem Zeitpunkt werden die Kerne des Spermiums und der Eizelle als männlicher bzw. weiblicher Vorkern bezeichnet. Die beiden Pronuklei nähern sich allmählich an, die Kernmembran verschwindet und die Chromosomen verschmelzen zu einer diploiden befruchteten Eizelle. Wenn die Spermien verschmolzen sind, kann man sehen, wie die Mikrovilli auf der Oberfläche der Eizelle die Spermien umgeben, die sich möglicherweise ausrichten; dann verschmilzt die Eierstockmembran mit der Plasmamembran im hinteren Bereich des Spermakrosoms.
Referenz
- Yanagimachi R. Germ cells and fertilization: why I studied these topics and what I learned along the path of my study. Andrology. 2014, 2(6):787-93.
- Henshaw J M, Marshall D J, Jennions M D, et al. Local gamete competition explains sex allocation and fertilization strategies in the sea. American Naturalist. 2014, 184(2):32.
- Mori T, Igawa T. Gamete attachment process revealed in flowering plant fertilization. Plant Signaling & Behavior. 2014, 9(12):977715.
- Hédouin L, Pilon R, Puisay A. Hyposalinitätsstress beeinträchtigt die Befruchtung von Gameten mehr als das Überleben von Korallenlarven. Marine Environmental Research. 2015, 104:1-9.
Luo J, Mcginnis L K, Carlton C, et al. PTK2b function during fertilization of the mouse oocyte. Biochem Biophys Res Commun. 2014, 450(3):1212-1217.