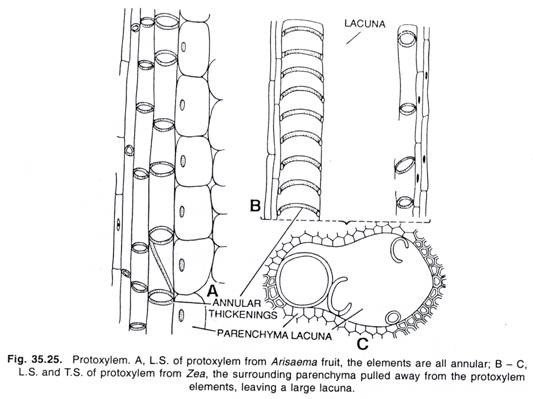
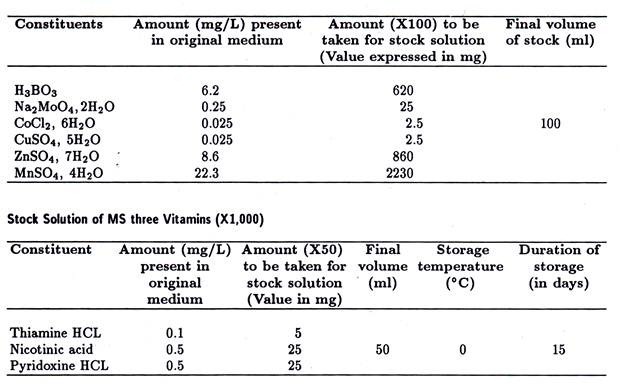
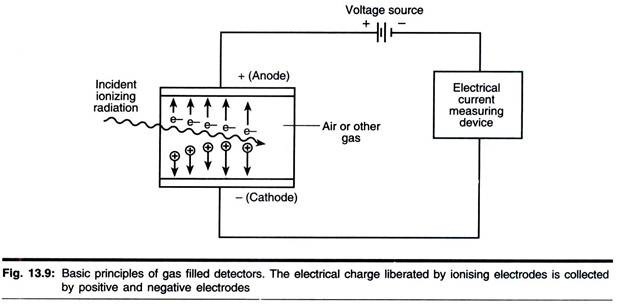
Hinweise:
In diesem Artikel werden wir über Paramecium Caudatum sprechen:- 1. Lebensraum, Habitat und Kultur von Paramecium Caudatum 2. Struktur von Paramecium Caudatum 3. Fortbewegung 4. Ernährung 5. Atmung und Ausscheidung 6. Osmoregulation 7. Verhalten 8. Fortpflanzung 9. Abweichendes Verhalten bei der Fortpflanzung 10. Einige zytoplasmatische Partikel.
Inhalt:
- Habitat, Lebensraum und Kultur von Paramecium Caudatum
- Struktur von Paramecium Caudatum
- Lokomotion von Paramecium Caudatum
- Ernährung von Paramecium Caudatum
- Respiration und Exkretion von Paramecium Caudatum
- Osmoregulation von Paramecium Caudatum
- Verhalten von Paramecium Caudatum
- Fortpflanzung in Paramecium Caudatum
- Aberrantes Verhalten bei der Fortpflanzung in Paramecium Caudatum
- Einige zytoplasmatische Partikel in Paramecium Caudatum
- Lebensraum, Lebensraum und Kultur von Paramecium Caudatum:
- Kultur von Paramecium:
- Struktur von Paramecium Caudatum:
- (i) Größe und Form von Paramecium Caudatum:
- (ii) Pellikel:
- (iii) Flimmerhärchen:
- (iv) Infra-Ciliaresystem:
- (v) Mundfurche und Cytopyge:
- (vi) Cytoplasma:
- (vii) Ektoplasma:
- (viii) Trichozysten:
- (ix) Neuromotorium und zugehörige Fibrillen:
- (x) Endoplasma:
- (xi) Kontraktile Vakuolen:
- (xii) Nahrungsvakuolen:
- Lokomotion von Paramecium Caudatum:
- (i) Metaboly oder Körperverrenkungen:
- (ii) Ciliäre Lokomotion:
- Ernährung von Paramecium Caudatum:
- Fütterungsmechanismus:
- Verdauung und Egestion von Paramecium Caudatum:
- Atmung und Ausscheidung von Paramecium Caudatum:
- Verhalten von Paramecium Caudatum:
- (i) Reaktionen auf Berührung (Thigmotaxis):
- (ii) Reaktionen auf Chemikalien (Chemotaxis):
- (iii) Reaktionen auf Temperatur (Thermotaxis):
- (iv) Reaktionen auf Licht (Phototaxis):
- (v) Reaktionen auf elektrischen Strom (Galvanotaxis):
- (vi) Reaktionen auf Wasserströmung (Rheotaxis):
- (vii) Reaktionen auf die Schwerkraft (Geotaxis):
- Fortpflanzung bei Paramecium Caudatam:
- (i) Transversale binäre Spaltung:
- (ii) Konjugation:
- Abweichendes Verhalten bei der Fortpflanzung in Paramecium Caudatum:
- (i) Endomixis:
- (ii) Autogamie:
- (iii) Hemixis:
- (iv) Zytogamie:
- Einige zytoplasmatische Partikel, die in Paramecium Caudatum gefunden wurden:
- (i) Kappa-Partikel:
- (ii) mµ, Partikel:
- (iii) Pi-Partikel:
- (iv) Lambda-Teilchen:
Lebensraum, Lebensraum und Kultur von Paramecium Caudatum:
Paramecium caudatum (Gr., paramekes = länglich; L., caudata = Schwanz) ist häufig in Süßwasserteichen, Tümpeln, Gräben, Bächen, Seen, Stauseen und Flüssen zu finden. Besonders häufig findet man sie in stehenden Teichen, die reich an Fäulnismaterial sind, in organischen Aufgüssen und im Abwasser. Paramecium caudatum ist ein freilebender Organismus und diese Art ist weltweit verbreitet.
Kultur von Paramecium:
Hinweise:
Nehmen Sie untergetauchtes Unkraut aus einem Teich und legen Sie es in ein Gefäß mit destilliertem Wasser, decken Sie das Gefäß ab und lassen Sie es verrotten; in wenigen Tagen werden Schwärme von Paramecien erscheinen. Nun koche Heu in Wasser, gieße den Aufguss ab, füge einige Weizenkörner hinzu und lasse ihn stehen, bis er von Bakterien getrübt ist.
Übertrage die Paramecien aus dem ersten Gefäß in diese Flüssigkeit, wo sie sich schnell vermehren werden. Heuinfusionen allein erzeugen Paramecien, die Zysten aufweisen, und es wurde von Zysten berichtet, die Sandkörnern ähneln, aber es gibt keinen Beweis dafür, dass Paramecien Zysten bilden, da sie nie bestätigt worden sind.
Struktur von Paramecium Caudatum:
(i) Größe und Form von Paramecium Caudatum:
Paramecium caudatum (Abb. 20.1) ist ein mikroskopischer Organismus und für das bloße Auge als winziger länglicher Körper sichtbar.
Hinweise:
Es erscheint hellgrau oder weiß und misst gewöhnlich zwischen 170 und 290 Mikrometer in der Länge und kann eine Länge von bis zu 300-350 Mikrometer erreichen. P. caudatum sieht aus wie die Sohle eines Pantoffels oder Schuhs, daher ist das Tier auch als Pantoffeltierchen bekannt. Es ist viermal so lang wie breit und etwas zylindrisch mit deutlich unterschiedlichen Enden.
Der sich vorwärts bewegende vordere Teil ist schlank und hat ein stumpfes oder abgerundetes Ende, während das hintere Ende etwas spitz oder kegelförmig ist. Die breiteste Stelle des Organismus befindet sich knapp unterhalb der Mitte. Der Körper des Tieres ist asymmetrisch geformt und weist eine gut ausgeprägte orale oder ventrale und eine aborale oder dorsale Oberfläche auf.
(ii) Pellikel:
Der Körper ist von einer dünnen, doppellagigen, elastischen und festen Pellicula aus Gelatine bedeckt. Die Pellicula hält die Form des Tieres, ist aber elastisch genug, um Kontraktionen zu ermöglichen. Die Pellikel hat eine doppelte Membran, wobei die äußere Membran mit den Flimmerhärchen und die innere Membran mit dem Ektoplasma verbunden ist. Unter der höheren Vergrößerung des Mikroskops zeigt die Pellikel rechteckige oder sechseckige Vertiefungen auf ihrer Oberfläche.

Diese Anordnung ist auf der dorsalen Oberfläche von Paramecium erhalten, aber auf der ventralen Oberfläche konvergieren die Kämme vorne und hinten zu einer präoralen und postoralen Apertur. Jede sechseckige Vertiefung ist von einer zentralen Öffnung durchbrochen, aus der ein einzelnes Cilium austritt. Die vorderen und hinteren Ränder der sechseckigen Vertiefungen tragen die Öffnungen der Trichocysten.
Die elektronenmikroskopische Untersuchung der Pellikel (Abb. 20.2) durch Ehret und Powers (1957) hat gezeigt, dass die hexagonalen Vertiefungen regelmäßigen Reihen von Hohlräumen, den Alveolen, entsprechen. Alle Alveolen zusammen bilden eine zusammenhängende Alveolarschicht, die durch eine äußere Alveolar- und eine innere Alveolarmembran abgegrenzt ist.
Die äußere Schicht liegt in engem Kontakt unter der äußeren Zellmembran. Daher umfasst die Pellikel die äußere Zellmembran, die äußere Alveolarmembran und die innere Alveolarmembran.
(iii) Flimmerhärchen:
Der gesamte Körper ist mit zahlreichen, kleinen, haarähnlichen Fortsätzen, den Cilien, bedeckt. Die Cilien treten in Längsreihen auf dem ganzen Körper auf, dieser Zustand wird als holotrich bezeichnet, bei dem die Körpercilien gleichmäßig angeordnet sind. Zilien haben die gleiche Struktur wie Geißeln, sie haben eine äußere Protoplasmahülle oder Plasmamembran mit neun doppelten Längsfibrillen in einem peripheren Ring. Bei einigen Zilien sind die neun äußeren Fibrillen nicht gepaart.
Hinweise:
Es gibt zwei zentrale Längsfibrillen, die dünner sind als die äußeren Fibrillen. Jedes Cilium entspringt aus einem Basalkörnchen oder Kinetosom. Die neun Paare peripherer Fibrillen verschmelzen miteinander, um die Wand des Kinetosoms zu bilden. Das Kinetosom ist also eine Röhre, die an ihrem unteren Ende entweder offen oder geschlossen ist; die beiden zentralen Fibrillen enden bei den meisten Ciliaten auf der Höhe der Pellikel.
Vom Kinetosom geht ein dünner Rhizoplast aus, der nicht mit dem Zellkern verbunden ist. Viele Metazoa haben auch Cilien, ihre Struktur ist die gleiche, außer dass das Basalkörnchen anders ist und feine Filamente oder Wurzelfasern hat, die sich in das Zytoplasma hinein erstrecken. Die Zilien unterscheiden sich jedoch von den Geißeln dadurch, dass sie im Allgemeinen zahlreicher und kürzer sind.
Die Flimmerhärchen können in Körper- oder somatische Flimmerhärchen, die sich auf der Körperoberfläche befinden, und in orale Flimmerhärchen, die mit der Mundregion verbunden sind, unterteilt werden. Die Körperwimpern sind gleich lang, aber am hinteren Ende sind sie länger, daher der Name caudatum. Die Flimmerhärchen sind Organellen der Fortbewegung und der Nahrungsaufnahme, sie fungieren auch als Sinnesrezeptoren und nehmen die Reize der äußeren Umgebung wahr.

Ultrastruktur der Cilien:
Die Flimmerhärchen und Geißeln sind fibrillär aufgebaut. An der Basis hat das Cilium einen Durchmesser von etwa 0,2 Mikron oder 2.000 A0, der bis zu 10 Mikron über der Zelloberfläche liegen kann. Die Flimmerhärchen werden von einer 90 A0 dicken Membran begrenzt, die der Plasmamembran ähnelt und mit ihr zusammenhängt. Der begrenzte Raum des Ciliums enthält eine wässrige Substanz, die als Matrix bezeichnet wird.
In die Matrix sind elf Längsfibrillen oder Mikrotubuli eingebettet. Von den elf Fibrillen befinden sich zwei in der Mitte, während die restlichen neun Fibrillen peripher um die zentralen Fibrillen herum angeordnet sind. Jede der neun äußeren Fibrillen hat einen Durchmesser von 360A0 und besteht aus zwei Unterfibrillen mit einem Durchmesser von 180 bis 250A0.
Diese Subfibrillen werden als Subfibrille A und Subfibrille B bezeichnet.
Hinweise:
Die Subfibrille A ist etwas größer als die Subfibrille B. Die Subfibrille A hat an einer Seite zwei dicke Fortsätze oder Arme. Die Arme der Subfibrille A aller äußeren Fibrillen bleiben im Uhrzeigersinn gerichtet. Außerdem liegt die Subfibrille A näher am Zentrum des Ciliums als die Subfibrille B. Beide Subfibrillen haben eine gemeinsame Wand von 50A0 Dicke.
Die beiden zentralen Fibrillen haben keine gepaarten Subfibrillen wie die neun peripheren Fibrillen, sondern enthalten jeweils nur ein einziges Röhrchen. Jede zentrale Fibrille hat einen Durchmesser von etwa 250A0 und besteht aus einer 60A0 dicken Wand.
Die beiden zentralen Fibrillen bleiben durch einen Abstand von 350A0 voneinander getrennt und sind von einer gemeinsamen Hülle umgeben. Gibbnos (1967) hat berichtet, dass die Hülle der zentralen Fibrillen neun radial ausgerichtete Glieder oder Speichen an jede Unterfibrille A abgibt.

Die hochauflösende Elektronenmikroskopie hat gezeigt, dass jede der peripheren und zentralen Fibrillen der Zilien und Geißeln aus zehn bis zwölf Filamenten von 40A0 Dicke besteht. Jedes Filament ist wulstig. Jede Perle ist in einem Gitter von 40 mal 50A0 in der Ebene der Röhrenwand angeordnet. Diese Wülste werden als die grundlegende Untereinheit der Röhrenstruktur betrachtet.
(iv) Infra-Ciliaresystem:
Das infraziliäre System befindet sich direkt unter den pellikulären Alveolen. Es besteht aus dem Kinetosom oder Basalkörper und dem Kinetodesma. Die Flimmerhärchen entstehen aus Kinetosomen, und aus jedem Kinetosom geht eine zarte zytoplasmatische Fibrille hervor, die Kinetodesma genannt wird (Abb. 20.2).
Unterhalb der Pellikel leicht rechts liegend, aber mit allen Kinetosomen einer Längsreihe verbunden, befindet sich ein Längsbündel aus mehreren Kinetodesmata, wobei ein Kinetodesmata jedes Kinetosoms ein Stück nach vorne in sein eigenes Kinetodesmabündel hineinragt.
Eine Längsreihe von Kinetosomen mit ihren Kinetodesmata bildet eine Längseinheit, die Kinetie genannt wird. Alle Kinetien oder Kinetien bilden das infraciliare System eines Ciliaten. Die Kinetien liegen in der Rinde unterhalb der Pellikel, ihre Anzahl ist bei jedem Wimpertierchen ziemlich konstant.
Das infraciliare System steuert und koordiniert die Bewegungen der Cilien, und es bewirkt die Bildung von Organellen bei der Zellteilung, z.B. bilden einige Kinetien den Mund. Bei der binären Spaltung der Wimpertierchen werden die Kinetien quer in zwei Teile geteilt, die jeweils in eine Tochterzelle gehen, dies wird als perikinetische Spaltung bezeichnet.
(v) Mundfurche und Cytopyge:
Auf der ventrolateralen Seite befindet sich eine große schräge, flache Vertiefung, die Mundfurche oder Peristom genannt wird und dem Tier ein asymmetrisches Aussehen verleiht. Sie verläuft schräg von einer Seite nach hinten (normalerweise von links nach rechts, in einigen Fällen aber auch von rechts nach links) und endet etwas hinter dem Mittelkörper. Die Mundfurche mündet in eine kurze konische, trichterförmige Vertiefung, die Vestibulum genannt wird.
Hinweise:
Das Vestibulum mündet direkt in die feste, ovale Öffnung, die Cytostom (Mund) genannt wird. Direkt vom Cytostom aus erstreckt sich der breite Cytopharynx in Richtung Körpermitte. Der Cytopharynx wendet sich dann scharf nach hinten und geht in den schlanken, spitz zulaufenden Ösophagus über.
Der Ösophagus verläuft also ungefähr parallel zur Körperoberfläche von Paramecium, außer an seinem hinteren Ende. Hier wendet sich die Speiseröhre wieder zur Mitte des Tieres, um in die sich bildende Nahrungsvakuole zu münden.
Die Cytopyge (auch Zellanus oder Analfleck oder Cytoprokt genannt) liegt an der ventralen Körperoberfläche fast senkrecht hinter dem Cytostom oder Mund. Unverdaute Nahrungspartikel werden durch die Cytopyge ausgeschieden. Die Ziliation des Cytopharynx ist sehr kompliziert.
Gelei (1934) berichtete über das Vorhandensein von vier Reihen und Lund (1941) beobachtete mindestens vier Reihen. An der linken Wand des Cytopharynx befindet sich eine Struktur namens Penniculus, die sich spiralförmig um etwa 90 Grad dreht, so dass ihr hinteres Ende auf der oralen (ventralen) Oberfläche des Ösophagus liegt.
Nach Lund besteht der Penniculus aus acht Reihen von Cilien, die in jeweils zwei eng beieinander liegenden Blöcken angeordnet sind. Ein ähnliches Band, das aus vier Reihen langer Flimmerhärchen besteht, die weniger kompakt sind als im Penniculus, wird als Quadrulus bezeichnet. Es zieht sich spiralförmig an der dorsalen Wand der Mundhöhle entlang und endet in der Nähe des Penniculus.
Der Penniculus und der Quadrulus wurden von einigen Arbeitern fälschlicherweise als eine wellenförmige Membran bezeichnet. Der Quadrulus und der Penniculus kontrollieren die Passage der Nahrung. Es ist nicht bekannt, wie die Flimmerhärchen funktionieren, wahrscheinlich ziehen sich ihre Fibrillen rhythmisch zusammen, was eine Biegung bewirkt. Gelei (1925) wies darauf hin, dass die Funktion des Penniculus darin besteht, die Nahrungsbestandteile in den Körper zu pressen.

(vi) Cytoplasma:
Das Cytoplasma wird in eine schmale, äußere oder kortikale Zone, Ektoplasma genannt, und eine größere, innere oder medulläre Region, Endoplasma genannt, unterschieden.
(vii) Ektoplasma:
Das Ektoplasma (Ektosark oder Kortex) ist ein fester Teil des Körpers, der sich deutlich vom Endoplasma abgrenzt. Das Ektoplasma bildet eine feste, klare, dünne und dichte äußere Schicht. Es enthält die Trichozysten, Zilien und fibrilläre Strukturen und ist nach außen durch eine Pellikel genannte Hülle begrenzt.
(viii) Trichozysten:
Im Ektoplasma sind rechtwinklig zur Oberfläche kleine spindelförmige Säckchen eingebettet, die Trichozysten genannt werden. Ein kleiner Punkt an jedem vorderen und hinteren Rand des Sechsecks markiert die Position einer Trichozyste. Sie sind mit einer brechenden, dichten Flüssigkeit gefüllt, die eine quellende Substanz enthält, am äußeren Ende befindet sich ein konischer Kopf oder Stachel.
Die Trichozysten liegen senkrecht im Ektoplasma, sie öffnen sich durch kleine Poren an den Graten der sechseckigen Bereiche der Pellikel.
Sie entstehen aus den Kinetosomen der Zilien, wandern dann und siedeln sich in gleicher Entfernung im Endoplasma an. Wenn das Tier gereizt wird, werden die Trichozysten als lange klebrige Fäden ausgeschieden. Eine ausgeschiedene Trichozyste hat einen undurchsichtigen Stachel, der wie ein umgedrehter Nagel aussieht, und einen langen, gestreiften Schaft, der jedoch im nicht ausgeschiedenen Zustand nicht zu sehen ist und wahrscheinlich während der Entladung gebildet wird.
Die Funktion der Trichozysten ist ungewiss, aber sie werden als Reaktion auf lokale Kontakte und Verletzungen ausgeschüttet, möglicherweise dienen sie als Abwehrorganellen.
Dies ist jedoch ungewiss, da die Trichozysten gegen Didinium, den Haupträuber von Paramecium, unwirksam sind; möglicherweise dienen sie dazu, das Tier während der Nahrungsaufnahme an einer Stelle zu fixieren. Bei einigen Wimpertierchen dienen die Trichozysten als Organellen des Angriffs. Nachdem die Trichozysten ausgeschieden sind, werden sie aus Kinetosomen regeneriert.


(ix) Neuromotorium und zugehörige Fibrillen:
Nach Lund (1933) befindet sich an der linken dorsalen Wand des Cytopharynx etwa in Höhe des hinteren Randes des Cytostoms eine sehr kleine, zweilappige Masse, das Neuromotorium. Vom Neuromotorium strahlen Fibrillen in das Endoplasma aus.
Von diesen reichen vier oder mehr normalerweise fast bis zur dorsalen Körperwand, aber die übrigen sind kürzer und in ihrer Lage nicht eindeutig. Alle werden als endoplasmatische Fibrillen bezeichnet. Ihre Funktion ist unbekannt, aber möglicherweise koordinieren sie die Fütterungsbewegungen der oralen Flimmerhärchen. Die Fibrillen können auch mechanische Unterstützung, Elastizität, Kontraktilität, Leitfähigkeit und metabolischen Einfluss geben.

(x) Endoplasma:
Das Endoplasma oder Medulla ist der flüssigere und voluminösere Teil des Zytoplasmas, der viele zytoplasmatische Körnchen sowie andere Einschlüsse und Strukturen spezialisierter Natur enthält. Zu den zytoplasmatischen Einschlüssen gehören Mitochondrien, Golgi-Apparate, Vakuolen, Kristalle, Granula und Chromidien usw. Andere Strukturen, nämlich Kerne, kontraktile Vakuolen und Nahrungsvakuolen, finden sich ebenfalls im Endoplasma.
Nuklei:
Im Endoplasma in der Nähe des Zytostoms befinden sich zwei Kerne, d.h. Paramecium ist heterokaryotisch, ein großer ellipsoidischer und körniger Makronukleus und ein weiterer kleiner kompakter Mikronukleus.
Der Makronukleus ist ein auffälliger, ellipsoider oder nierenförmiger Körper. Er ist vom kompakten Typ, enthält feine Fäden und dicht gepackte diskrete Chromatinkörnchen unterschiedlicher Größe und ist in eine achromatische Matrix eingebettet. Es besitzt viele Nukleoli und viel mehr Chromatinmaterial (DNA).
Es ist ein somatischer oder vegetativer Zellkern. Er teilt sich amitotisch und kontrolliert die vegetativen Funktionen (Stoffwechselaktivitäten) des Tieres. Er macht eine Mitose durch.
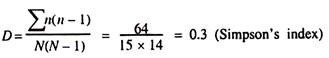
Der Mikronukleus ist klein, kompakt und kugelig. Er befindet sich im Allgemeinen in der Nähe des Makronucleus, oft in einer Konkavität. Feine Chromatinkörnchen und -fäden sind gleichmäßig in der Struktur verteilt. Der Mikronukleus teilt sich mitotisch und kontrolliert die Fortpflanzung.
Die Anzahl der Mikronuklei variiert je nach Art; sie beträgt einen bei P. caudatum, zwei bei P. aurelia und viele bei P. multimicronucleatum. Der Mikronukleus enthält bei P. aurelia einen ausgeprägten Nukleolus, der bei P. caudatum nicht vorhanden ist. Moses (1949; 1950) hat berichtet, dass der Makronukleus und der Mikronukleus in ihrer chemischen Zusammensetzung identisch sind.
(xi) Kontraktile Vakuolen:
Es gibt zwei große, flüssigkeitsgefüllte kontraktile Vakuolen, die sich jeweils in der Nähe eines Endes des Körpers nahe der dorsalen Oberfläche befinden. Ihre Position ist fest (im Gegensatz zu Amöben), sie liegen zwischen dem Ektoplasma und dem Endoplasma, aber sie sind temporäre Organellen, die periodisch verschwinden. Bei einigen Arten scheinen sie eine Auskleidungsmembran zu haben, in diesem Fall verschwinden sie während der Systole nicht vollständig.
Mit jeder kontraktilen Vakuole sind fünf bis zwölf röhrenförmige Strahlenkanäle verbunden, die jeweils aus einem Endteil, einer langen Ampulle, die zusammenfällt, wenn sie leer ist, und einem kurzen Injektorkanal bestehen, der sich in die Vakuole öffnet.

Die Kanäle stehen mit einem großen Teil des Körpers in Verbindung, von wo aus sie Flüssigkeiten aufnehmen und in die Vakuole gießen, die so wiederhergestellt wird und an Größe zunimmt; wenn die kontraktile Vakuole ihre maximale Größe erreicht, zieht sie sich plötzlich zusammen (Systole) und entlädt ihren Inhalt durch eine permanente Pore in der Pellikel, dann bilden die Kanäle wieder die kontraktilen Vakuolen, die Kanäle verschwinden nicht ganz, da sie permanente Strukturen sind.
Die beiden kontraktilen Vakuolen entleeren sich unregelmäßig, wobei sich die hintere schneller zusammenzieht, weil sie sich in der Nähe des Cytopharynx befindet und mehr Wasser in sie eindringt. Die Hauptfunktion der Kanäle und der kontraktilen Vakuolen ist hydrostatisch, sie entziehen dem Protoplasma überschüssiges Wasser, das bei der Nahrungsaufnahme teilweise absorbiert und teilweise aufgenommen wird.
Die stickstoffhaltigen Abfallstoffe bestehen aus Ammoniakverbindungen und einigen Uraten, die zusammen mit CO2 aus den kontraktilen Vakuolen ausgeschieden werden, aber es gibt keinen Hinweis darauf, dass die Ausscheidungsstoffe vom Protoplasma in die Kanäle oder kontraktilen Vakuolen ausgeschieden werden.
Unterhalb des Peristoms befindet sich auf einer Seite des Cytopharynx ein fester permanenter Cytopyge oder Anus, durch den unverdaute Nahrungsreste und Urate ausgeschieden werden.
Die elektronenmikroskopische Untersuchung der kontraktilen Vakuolen hat ergeben, dass jeder kontraktile Apparat aus einigen Tubuli des endoplasmatischen Retikulums, nephridialen Tubuli, Zubringerkanälen, akzessorischen Vakuolen (Radialkanälen) und der kontraktilen Hauptvakuole besteht. Bei den akzessorischen Vakuolen handelt es sich vermutlich um die Ampullen der Zubringerkanäle (Abb. 20.11).
(xii) Nahrungsvakuolen:
Es handelt sich um etwa kugelförmige, nicht kontraktile Körper, die in Größe und Anzahl variieren und im Endoplasma liegen. Sie enthalten aufgenommene Nahrungspartikel, vor allem Bakterien und eine kleine Menge Flüssigkeit, die von einer dünnen, festen Membran begrenzt wird. Volkonsky (1934) schlug für diese Vakuolen den Namen Gastriolen vor. Mit den Nahrungsvakuolen assoziiert sind die Verdauungsgranula.
Lokomotion von Paramecium Caudatum:
Paramecium Caudatum führt die Fortbewegung auf zwei Arten durch, nämlich durch metabole oder körperliche Verrenkungen und durch Flimmerhärchen.
(i) Metaboly oder Körperverrenkungen:
Der Körper von Paramecium Caudatum besitzt Elastizität, er kann sich durch einen Durchgang quetschen, der enger ist als sein Körper, woraufhin der Körper seine normale Form annimmt. Diese vorübergehende Veränderung der Körperform ist metaboly, sie wird bei Paramecium durch das Protoplasma bewirkt.
(ii) Ciliäre Lokomotion:
Die Hauptmethode ist die durch Zilien bewirkte Lokomotion. Die Flimmerhärchen können vorwärts oder rückwärts schlagen, so dass das Tier nach vorne oder nach hinten schwimmen kann.
Normalerweise schwimmt das Tier vorwärts, die Flimmerhärchen schlagen nach hinten, aber schräg, die Flimmerhärchen versteifen sich und biegen sich schnell nach hinten, um fast die Körperoberfläche zu berühren, dies wird als effektiver Schlag bezeichnet; dann werden die Flimmerhärchen schlaff und kehren langsam in die ursprüngliche vertikale Position zurück, dies wird als Erholungsschlag bezeichnet.
Die Flimmerhärchen der gleichen Querreihe schlagen zusammen und die der gleichen Längsreihe schlagen nacheinander vom vorderen zum hinteren Ende.

Diese koordinierte Bewegung der Flimmerhärchen wird metachroner Rhythmus genannt, der auf das infraciliare System zurückzuführen ist; dies bewirkt, dass das Tier vorwärts schwimmt. Wenn aber die Körpercilien schräg nach hinten schlagen, dann schlagen gleichzeitig die längeren Cilien der Mundrinne kräftiger, was das vordere Ende nach links ausschlagen lässt.
Die Wirkung der Flimmerhärchen des Körpers und der Mundrinne bewirkt, dass sich das Tier um seine Längsachse dreht. Diese Drehung ist immer nach links gerichtet (außer bei P. calkinsi, die sich in einer Rechtsspirale dreht).
Durch die Kombination von Vorwärtsbewegung, Ausweichen und Drehung bewegt sich das Tier auf einer spiralförmigen Bahn gegen den Uhrzeigersinn vorwärts. Diese Bahn hat eine gerade Achse, und die gleiche Körperoberfläche des Tieres bleibt in Richtung der Achse der Spiralbahn. Aber beim Rückwärtsschwimmen drehen sich alle Arten nach rechts.
Der Zilienschlag kann umgekehrt werden, so dass sich die Zilien schräg nach vorne bewegen, wodurch das Tier rückwärts schwimmt. Durch die Zilientätigkeit bewegt sich Paramecium mit einer Geschwindigkeit von 1500 Mikrometern oder sogar mehr pro Sekunde.
Jennings vermutet, dass die spiralförmige Bewegung von Paramecium darauf zurückzuführen ist, dass die Cilien zwar hauptsächlich nach hinten schlagen, dies aber schräg nach rechts tun, wodurch sich das Tier nach links dreht.
Auch dieses Ausweichen des Körpers in Richtung der aboralen Oberfläche ist größtenteils auf die größere Kraft des effektiven Schlags der oralen Flimmerhärchen zurückzuführen, die direkter nach hinten schlagen. Das Ergebnis – die Drehung des Parameciums um seine Längsachse – ermöglicht es dem Paramecium, bei der Bildung großer Spiralen einen mehr oder weniger geraden Verlauf zu nehmen.
Ernährung von Paramecium Caudatum:
In Paramecium Caudatum ist die Ernährung holozoisch. Die Nahrung besteht hauptsächlich aus Bakterien und winzigen Protozoen. Paramecium wartet nicht auf die Nahrung, sondern jagt aktiv nach ihr.
Es wird behauptet, dass Paramecium Caudatum bei der Auswahl seiner Nahrung wählerisch ist, aber dafür scheint es keine Grundlage zu geben, obwohl es nur bestimmte Arten von Bakterien verschlingt; verfügbare Daten deuten darauf hin, dass 2 bis 5 Millionen Individuen von Bacillus coli von einem einzigen Paramecium in 24 Stunden verschlungen werden. Es ernährt sich auch von einzelligen Pflanzen wie Algen, Kieselalgen usw. sowie von kleinen Tier- und Gemüsestücken.
Fütterungsmechanismus:
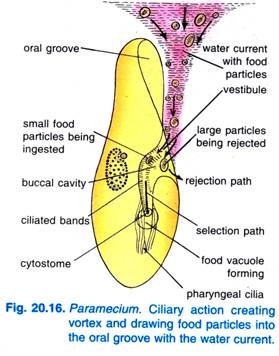
Wenn Paramecium Caudatum in eine Region mit reichlich Nahrung kommt, ruht es sich aus. Es ernährt sich nur in Ruhe oder wenn es sehr langsam schwimmt, es ernährt sich nie, wenn es schnell schwimmt. Das Schlagen der Flimmerhärchen in der Mundrinne bewirkt, dass ein kegelförmiger Wirbel mit nahrungshaltigem Wasser aus einer Entfernung vor dem vorderen Ende in die Mundrinne gespült wird (Abb. 20.16).
Die Nahrungspartikel gelangen dann in das Vestibulum, von wo aus einige Nahrungspartikel abgestoßen und ausgeschleudert werden, andere aber in das Zytostom gelangen.
Am Ende des Cytopharynx bildet sich eine Nahrungsvakuole, die mit Nahrungspartikeln gefüllt wird. Der Quadrulus und Peniculi kontrollieren den Übergang der Nahrung in die seitlich gebildete Nahrungsvakuole. Wenn die Nahrungsvakuole eine bestimmte Größe erreicht hat, umklammern die post-bukkalen Fasern die Nahrungsvakuole und sie wird von ihnen abgeklemmt und setzt ihren Weg fort.
Die Vakuole enthält neben der Nahrung auch etwas Wasser. Rotierende, strömende Bewegungen des Endoplasmas, die Zyklose genannt werden, tragen die Nahrungsvakuole entlang eines bestimmten Kurses, der funktionell einem Verdauungstrakt entspricht.
Der Trakt beginnt am Ende des Cytopharynx, dann zur hinteren Seite, dann nach vorne, um mit dem Endoplasma zu zirkulieren, dann zur dorsalen Oberfläche, dann zum vorderen Ende, dann nach unten zur Cytopyge. Zu Beginn ihrer Reise nimmt die Nahrungsvakuole an Größe ab, dann nimmt sie wieder zu.

Verdauung und Egestion von Paramecium Caudatum:
Während der Zyklose erfolgt die Verdauung durch Enzyme, die vom Protoplasma in die Vakuolen ausgeschieden werden. Bei der Verdauung werden Proteine in Aminosäuren, Kohlenhydrate in lösliche Zucker und Glykogen umgewandelt, und wahrscheinlich werden auch Fette verdaut.
Der Inhalt der Nahrungsvakuolen ist zunächst sauer (pH-Wert etwa 4) und wird dann alkalisch, wobei die Hauptverdauung während der alkalischen Phase stattfindet. Die unverdaute Materie wird mit einiger Kraft durch die Zytopyge ausgestoßen.
Die Zyklose kann experimentell nachgewiesen werden; wenn mit Kongorot gefärbte Milch an Paramecium verfüttert wird, färben sich die Fettkügelchen der Milch in den Nahrungsvakuolen aufgrund der sauren Reaktion der Enzyme zunächst rot, dann wechseln sie aufgrund der alkalischen Reaktion von violetten zu blauen Farbtönen; die Vakuolen zeigen den Verlauf der Zyklose.
Atmung und Ausscheidung von Paramecium Caudatum:
Der Austausch von Gasen (Sauerstoff und Kohlendioxid) erfolgt durch die semipermeable Pellikel wie bei anderen Süßwasserprotozoen durch den Prozess der Diffusion. Paramecium Caudatum bezieht seinen Sauerstoff aus dem umgebenden Wasser. Kohlendioxid und organische Abfälle wie Ammoniak aus dem Stoffwechsel werden wahrscheinlich durch Diffusion in umgekehrter Richtung nach außen ins Wasser ausgeschieden.

6. Osmoregulation in Paramecium Caudatum:
Paramecium Caudatum hat zwei kontraktile Vakuolen, eine anteriore und eine posteriore. Die Funktion der kontraktilen Vakuolen ist die Osmoregulation, d.h. die Regulierung des Wassergehalts des Körpers und kann auch der Ausscheidung von stickstoffhaltigen Abfallstoffen wie Harnstoff und Ammoniak dienen.
Überschüssiges Wasser (aufgrund kontinuierlicher Endosmose) im Zytoplasma wird in die Tubuli des endoplasmatischen Retikulums abgesondert und gelangt in die Nephridialtubuli → Zubringerkanäle → und sammelt sich in der Ampulla einer Reihe von 6 bis 11 strahlenförmig verlaufenden Kanälen, die zu jeder Vakuole konvergieren und in diese münden. Die Kanäle sind am auffälligsten, wenn sich eine Vakuole bildet.
Wenn jede Vakuole bis zu einer bestimmten Größe angeschwollen ist (Diastole), zieht sie sich zusammen (Systole) und entlädt sich wahrscheinlich durch eine Pore nach außen. Die kontraktilen Vakuolen kontrahieren abwechselnd, in Abständen von 10 – 20 Sekunden.
Die hintere kontraktile Vakuole arbeitet schneller als die vordere Vakuole, da der Cytopharynx eine große Menge Wasser in die hintere Region aufnimmt. Die kontraktilen Vakuolen halten eine optimale Wasserkonzentration im Körperzytoplasma aufrecht, indem sie den Überschuss entsorgen.

Verhalten von Paramecium Caudatum:
Die Reaktionen von Paramecium Caudatum auf verschiedene Arten von Reizen werden durch das Studium seiner Reaktionen und der Gruppierung oder Streuung von Individuen in einer Kultur erlernt. Die Reaktion ist positiv, wenn sich das Tier auf einen Reiz zubewegt, und negativ, wenn es sich davon entfernt. Auf einen ungünstigen Reiz reagiert das Tier so lange ausweichend, bis es entkommt.
Bei der Ausweichreaktion kehrt sich der Ziliarschlag um, das Tier bewegt sich eine kurze Strecke rückwärts und dreht sich dann auf einer konischen Bahn, indem es das vordere Ende nach aboral schwenkt, während es sich auf der hinteren Spitze dreht. Alle Anpassungen werden durch Versuch und Irrtum vorgenommen. Experimente haben gezeigt, dass das vordere Ende des Tieres empfindlicher ist als die anderen Teile.
Die Reaktionen von Paramecium auf verschiedene Reize können wie folgt gruppiert werden:
(i) Reaktionen auf Berührung (Thigmotaxis):
Die Reaktion auf Berührung ist bei Paramecium unterschiedlich. Wenn das vordere Ende mit einer feinen Spitze leicht berührt wird, kommt es zu einer starken Ausweichreaktion. Wenn ein schwimmendes Paramecium mit einem Objekt im Wasser kollidiert, kann es sein, dass es keine Reaktion zeigt, wenn es an anderer Stelle berührt wird. Ein sich langsam bewegendes Individuum reagiert oft positiv auf die Berührung eines Objekts, indem es auf diesem zur Ruhe kommt.
(ii) Reaktionen auf Chemikalien (Chemotaxis):
Generell reagieren Paramecien auf chemische Reize mit einer Vermeidungsreaktion. Wird ein Tropfen einer schwachen Salzlösung (0,5 Prozent) in eine Paramecium-Population auf einem Mikroschlitten eingebracht, reagieren die Tiere mit der Ausweichreaktion und keines tritt in den Tropfen ein. Auf Säuren hingegen ist die Reaktion positiv, selbst wenn die Konzentration stark genug ist, um sie zu töten.
(iii) Reaktionen auf Temperatur (Thermotaxis):
Paramecium strebt eine optimale Temperatur von 24 bis 28°C an. Bei einer Temperaturänderung, die deutlich über oder unter dem optimalen Bereich liegt, zeigen Paramecien eine Vermeidungsreaktion. Größere Hitze stimuliert schnelle Bewegungen und Vermeidungsreaktionen, bis die Tiere entkommen oder getötet werden.
(iv) Reaktionen auf Licht (Phototaxis):
Mit Ausnahme des grünen Paramecium bursaria, das positiv phototaktisch ist, sind die anderen Arten gegenüber gewöhnlichem Licht gleichgültig. Wenn jedoch die Lichtintensität plötzlich und stark erhöht wird, folgt im Allgemeinen eine negative Reaktion. Paramecien zeigen eine sofortige negative Reaktion auf ultraviolette Strahlen.
(v) Reaktionen auf elektrischen Strom (Galvanotaxis):
Paramecien reagieren auf elektrische Reize. Wenn zwei Elektroden in einer flachen Schale mit Paramecien einander gegenüberliegen und ein konstanter Strom angelegt wird, schwimmen alle Organismen in die gleiche Richtung zur Kathode oder negativen Elektrode, wo sie sich in großer Zahl konzentrieren.
Wenn die Richtung des elektrischen Stroms umgekehrt wird, während die Paramecien zur Kathode schwimmen, kehren die Organismen die Richtung um und schwimmen zur neuen Kathode.

(vi) Reaktionen auf Wasserströmung (Rheotaxis):
Paramecien zeigen eine positive Rheotaxis. In einer leichten Wasserströmung bewegen sich die Paramecien meist mit der Strömung mit ihren vorderen Enden stromaufwärts.
(vii) Reaktionen auf die Schwerkraft (Geotaxis):
Paramecien reagieren im Allgemeinen negativ auf die Schwerkraft, wie man in einer Kultur sehen kann, in der sich viele Individuen dicht unter dem Oberflächenfilm sammeln, wobei ihre vorderen Enden nach oben gerichtet sind. Werden Paramecien in ein umgedrehtes, mit Wasser gefülltes und an beiden Enden verschlossenes U-förmiges Rohr eingebracht, so bewegen sie sich sofort nach oben in den horizontalen Teil des Rohrs.
Fortpflanzung bei Paramecium Caudatam:
Paramecium Caudatum pflanzt sich ungeschlechtlich durch transversale Binärspaltung fort und durchläuft außerdem mehrere Arten von Kernumbau, wie Konjugation, Endomixis, Autogamie, Cytogamie und Hemixis, usw.
(i) Transversale binäre Spaltung:
Die transversale Binärspaltung ist die häufigste Art der asexuellen Fortpflanzung bei Paramecium. Es handelt sich um einen einzigartigen asexuellen Prozess, bei dem sich ein ausgewachsenes Exemplar in zwei Tochterindividuen teilt, ohne einen elterlichen Körper zu hinterlassen.
Die Teilungsebene verläuft durch das Zentrum der Zelle und in einer Ebene, die rechtwinklig zur Längsachse des Körpers verläuft. Der Teilung des Zellkörpers als Ganzes geht immer eine Teilung der Kerne voraus; es scheint sogar, dass die Fortpflanzung durch Kernaktivität und -teilung eingeleitet wird.

Paramecium Caudatum vermehrt sich unter günstigen Bedingungen durch transversale binäre Spaltung. Bei der binären Spaltung teilt sich der Mikrokern durch Mitose in zwei Tochtermikrokerne, die sich an entgegengesetzte Enden der Zelle bewegen. Der Makronukleus dehnt sich aus und teilt sich durch Amitose in Querrichtung.
Ein weiterer Cytopharynx wird abgespalten und zwei neue kontraktile Vakuolen entstehen, eine am vorderen und eine am hinteren Ende. In der Zwischenzeit erscheint eine Einschnürungsfurche nahe der Körpermitte und vertieft sich, bis das Zytoplasma vollständig geteilt ist.
Die daraus entstehenden zwei „Tochter“-Paramecien sind gleich groß und enthalten jeweils einen Satz von Zellorganellen. Von den beiden entstehenden Tochterparamecien wird die vordere als Proter und die hintere als Opisthe bezeichnet. Sie wachsen zu voller Größe heran, bevor eine weitere Teilung stattfindet.
Der Prozess der binären Spaltung dauert etwa zwei Stunden und kann ein- bis viermal am Tag stattfinden, wobei 2 bis 16 Individuen entstehen. In einem Jahr werden etwa 600 Generationen erzeugt.
Die Vermehrungsrate hängt von den äußeren Bedingungen wie Nahrung, Temperatur, Alter der Kultur und Bevölkerungsdichte ab, aber auch von den inneren Faktoren wie Vererbung und Physiologie. Würden alle Nachkommen eines Individuums überleben und sich fortpflanzen, so würde die Zahl der produzierten Paramecien natürlich bald dem Volumen der Erde entsprechen.
Als Klon bezeichnet man alle Individuen, die aus einem Individuum durch Spaltung hervorgegangen sind. Alle Mitglieder eines Klons sind erblich gleich.
(ii) Konjugation:
Ordinär vermehrt sich Paramecium Caudatum durch binäre Spaltung über lange Zeiträume, aber in Intervallen kann dies durch die Vereinigung zweier Tiere entlang ihrer Mundflächen für den sexuellen Prozess der Konjugation unterbrochen werden.
Konjugation ist definiert als die vorübergehende Vereinigung zweier Individuen, die gegenseitig Mikrokernmaterial austauschen. Es handelt sich um einen einzigartigen Typ eines sexuellen Prozesses, bei dem sich zwei Organismen bald nach dem Austausch des Kernmaterials trennen.
Sonneborn (1947) hat auf der Grundlage des Paarungsverhaltens von Paramecium Caudatum berichtet, dass jede Art von Paramecium in einer Reihe von Varietäten oder Syngen existiert. Außerdem gibt es innerhalb jedes Syngens eine Anzahl von Paarungstypen, gewöhnlich zwei.
Die Paarungstypen bleiben morphologisch identisch, weisen aber physiologische Unterschiede auf. Bei P. aurelia gibt es 14 Syngen und 28 Paarungstypen, während es bei P. caudatum 16 Syngen und 32 Paarungstypen gibt. Es wurde beobachtet, dass Paramecien in der Regel weder mit Mitgliedern ihres eigenen Paarungstyps noch mit den anderen Varietäten konjugieren, sondern nur mit dem zweiten Paarungstyp ihrer eigenen Varietät.

Faktoren, die die Konjugation induzieren:
Die Faktoren, die die Konjugation auslösen, variieren von Spezies zu Spezies, aber einige von ihnen sind unten aufgeführt:
1. Die Konjugation tritt gewöhnlich unter ungünstigen Lebensbedingungen auf; Hunger oder Nahrungsmangel und eine besondere bakterielle Ernährung oder bestimmte Chemikalien sollen den Prozess der Konjugation bei bestimmten Arten von Paramecium auslösen.
2. die Konjugation tritt nach etwa 300 asexuellen Generationen der binären Spaltung auf, oder sie wechselt mit der binären Spaltung in langen Abständen ab, um den sterbenden Klon zu verjüngen, d.h. sie tritt bei den Individuen auf, die eine wünschenswerte Anzahl asexueller Generationen durchlaufen haben müssen, was als Periode der Unreife bezeichnet wird, und dann werden sie geschlechtsreif, um zu konjugieren.
3. die Konjugation findet statt, wenn sich der physiologische Zustand der Paramecien ändert, dann findet sie zwischen solchen Individuen statt, die etwas kleiner sind (210 Mikron lang) und sich in einem Stadium befinden, das als eine Periode ungesunden Alters angesehen werden kann; die Paramecien in diesem Zustand sterben, wenn man ihnen nicht erlaubt, sich zu konjugieren.
4. Plötzliche Dunkelheit in den Lichtverhältnissen und niedrige Temperaturen sollen bei einigen Arten den Prozess der Konjugation auslösen.
5. Die Konjugation findet nicht während der Nacht oder Dunkelheit statt; sie beginnt am frühen Morgen und dauert bis zum Nachmittag.
6. Eine eiweißhaltige Substanz in den Zilien von Individuen des Paarungstyps soll die Konjugation auslösen.
Vorgang der Konjugation:
Der Vorgang der Konjugation ist bei den verschiedenen Paramecium-Arten unterschiedlich, doch bezieht sich die unterstehende Darstellung auf den Konjugationsvorgang von P. caudatum (Abb. 20.21).
Bei der Konjugation kommen zwei Paramecium caudatum (als Vorkonjugierte bezeichnet) der entgegengesetzten Paarungstypen der gleichen Art mit ihren Ventralflächen zusammen und vereinigen sich durch ihre Mundrillen; ihre Cilien produzieren eine Substanz auf der Körperoberfläche, die eine Adhäsion der beiden konjugierenden Paramecien bewirkt.
Sie stellen die Nahrungsaufnahme ein und ihr Mundrillenapparat verschwindet. Die Pellikel und das Ektoplasma brechen an der Berührungsstelle der beiden auf, und es bildet sich eine protoplasmatische Brücke zwischen den beiden Tieren. Diese Individuen werden nun Konjuganten genannt.
In diesem Zustand schwimmt das konjugierende Paar aktiv und gleichzeitig findet in jedem Konjuganten eine Reihe von Kernveränderungen statt, die im Folgenden beschrieben werden:
Der Makronukleus beginnt sich aufzulösen, er wird locker und bildet ein komplexes, verdrehtes Geflecht; in der zweiten Hälfte der Konjugationsperiode verschwindet er schließlich, indem er im Zytoplasma absorbiert wird. Der Mikronukleus jeder Konjugation teilt sich zweimal, eine davon ist eine Reduktionsteilung.
Auf diese Weise entstehen in jeder Konjugation vier haploide Tochtermikrokerne. Drei dieser vier Mikronuklei degenerieren in jedem, so dass nur einer übrig bleibt.
Der verbleibende Mikronukleus jeder Konjugation teilt sich mitotisch in zwei ungleiche Pronuklei oder gametische Kerne, die einen größeren stationären weiblichen Pro-Kern und einen kleineren, aktiven wandernden männlichen Pro-Kern bilden.
Der wandernde Pro-Kern des einen Konjuganden überquert die protoplasmatische Brücke und verschmilzt mit dem stationären Pro-Kern des anderen Konjuganden, um ein Synkaryon oder einen Konjugationskern zu bilden, in dem die diploide Chromosomenzahl wiederhergestellt ist und ein Austausch des Erbmaterials stattgefunden hat.
Der Vorgang wurde mit der Befruchtung bei höheren Tieren verglichen, aber es handelt sich nicht um eine Befruchtung, da keine Gameten beteiligt sind. Die Konjuganten trennen sich nun (nach etwa 12-48 Stunden) und werden Ex-Konjuganten genannt. Das Synkaryon jedes Ex-Konjuganten teilt sich dreimal und bildet acht Mikrokerne in jedem Ex-Konjuganten.
Vier der acht Mikronuklei vergrößern sich und werden zu Makronuklei, und drei der anderen vier Mikronuklei verschwinden.
Der verbleibende Mikronukleus teilt sich und gleichzeitig teilt sich der Ex-Konjugant durch binäre Spaltung in zwei Zellen, die jeweils zwei Makronuklei und einen Mikronukleus haben. Die Zellen und ihre Mikrokerne teilen sich ein zweites Mal, um aus jedem Ex-Konjuganten vier Paramecien zu bilden, so dass jede einen Makronukleus und einen Mikrokern hat.
Der neue Makronukleus, wie auch der Mikronukleus, sind aus neuem Material hergestellt worden. Diese neuen Kerne enthalten wahrscheinlich ein neues und anderes Potential, das sich in den gesunden Individuen widerspiegelt.
Bedeutung der Konjugation:
Ein Klon stirbt aus, wenn keine nukleare Reorganisation stattfindet, aber der Klon kann verjüngt werden, um seine frühere Vitalität durch nukleare Reorganisation wiederzuerlangen; diese nukleare Reorganisation wird durch Konjugation herbeigeführt, daher ist die Konjugation für eine fortgesetzte Binärspaltung unerlässlich.
Die Bedeutung der Konjugation wird im Folgenden zusammengefasst:
1. Die Konjugation dient als Prozess der Verjüngung und Reorganisation, durch den die Vitalität der Rasse wiederhergestellt wird. Wenn die Konjugation über längere Zeiträume nicht stattfindet, werden die Paramecien geschwächt und sterben ab. (Woodruffs Behauptung, Paramecien über 22.000 Generationen ohne Konjugation gesund erhalten zu haben, wird von Sonneborn widerlegt, weil er zeigte, dass alle Woodruffschen Paramecien demselben Paarungstyp angehörten).
2. es gibt keinen Geschlechtsunterschied bei den Konjuganten, obwohl nur Paramecien von zwei verschiedenen Paarungstypen derselben Sorte konjugieren können.
3. Es gibt keine Unterscheidung des Geschlechts, aber der aktive wandernde Vorkern wird als männlich und der stationäre Vorkern als weiblich angesehen.
4. die Konjugation ist nur eine vorübergehende Vereinigung, es findet keine Verschmelzung des Zytoplasmas statt und es wird keine Zygote erzeugt, aber der Kern jedes Ex-Konjuganden enthält Erbmaterial von zwei konjugierenden Individuen.
5. Bei der Konjugation wird der Makronukleus durch Material aus dem Synkaryon ersetzt, was ein Ereignis von grundlegender Bedeutung ist. Bei der binären Spaltung wurden die Chromosomen des Makronukleus nach dem Zufallsprinzip auf die Tochterzellen verteilt, die fortgesetzte binäre Spaltung hatte den Klon schwach gemacht mit einigen strukturellen Anomalien.
Die Konjugation bewirkt die Bildung der korrekten Anzahl von Chromosomen im Makronukleus, so dass die Rasse in ihrer Vitalität erneuert wird. Die Aufgabe des Mikrokerns besteht darin, das Gleichgewicht des Chromosomen- und Genkomplexes wiederherzustellen.
Abweichendes Verhalten bei der Fortpflanzung in Paramecium Caudatum:
Paramecium Caudatum zeigt gewisse Abweichungen in seinem Kernverhalten während der Spaltung und Konjugation, diese Abweichungen sind Endomixis, Autogamie, Cytogamie und Hemixis. Bei den ersten drei Prozessen findet eine genetische Rekombination statt und aus dem Mikrokern wird ein neuer Makrokern gebildet.
(i) Endomixis:
Woodruff und Erdmann (1914) berichteten zuerst über einen neuen Prozess der Kernreorganisation, die Endomixis (gr., endon = innerhalb; mixis = Vermischung) bei Paramecium aurelia, einer zweikeimblättrigen Art (Abb. 20.22). Dieser Prozess wurde als periodisch auftretender Prozess beschrieben, bei dem ein neuer makrokerniger Apparat ohne Synkaryonbildung produziert wird. Die Endomixis tritt innerhalb eines einzigen Individuums auf.
Nach Woodruff und Erdmann degeneriert der Makronukleus und die Mikronuklei teilen sich zweimal und bilden acht Mikronuklei. Sechs Mikrokerne degenerieren und zwei bleiben übrig. Mit nur noch zwei Mikrokernen teilt sich das Tier durch Spaltung in zwei Zellen mit je einem Mikrokern.
Der Mikronukleus jeder Tochterzelle teilt sich zweimal und bildet vier Mikronuklei. Zwei Mikronuklei vergrößern sich zu Makronuklei. Das Tier und seine Mikrokerne teilen sich, so dass zwei Tochterindividuen entstehen, die jeweils einen Makronukleus und zwei Mikrokerne haben.
Die Endomixis tritt bei der Sorte von P. aurelia auf, die nicht konjugiert, daher kann die Wirkung der Endomixis die gleiche sein wie die der Konjugation, da beide Prozesse den Ersatz des Makronukleus durch Material aus dem Mikronukleus bewirken und beide Prozesse die Vitalität der Rasse verjüngen.
Die beiden Prozesse unterscheiden sich jedoch dadurch, dass bei der Endomixis keine Verschmelzung der Vorkerne stattfindet; die Endomixis kann mit der Parthenogenese verglichen werden.
Einige Forscher haben jedoch mit guten Gründen behauptet, dass die Endomixis kein gültiger Prozess ist und dass sie aufgrund fehlerhafter Beobachtungen beschrieben wurde. Höchstwahrscheinlich findet Endomixis nicht statt und ist möglicherweise nur ein Spezialfall der Autogamie.

Später berichteten Erdmamm und Woodruff (1916) über Endomixis bei Paramecium caudatum. Diller glaubt jedoch nicht an die Gültigkeit dieses Prozesses und ist der Meinung, dass Erdmann und Woodruff einfach die Stadien der Hemixis und Autogamie zu einem Schema, der Endomixis, zusammengefasst haben.
(ii) Autogamie:
Diller (1934, 1936) und Sonneborn (1950) beschrieben einen Prozess der Selbstbefruchtung oder Autogamie, der bei einem einzigen Individuum in Paramecium aurelia auftritt (Abb. 20.23). Er berichtete, dass bei der Autogamie drei mikrokernige (prägamische) Teilungen, die eine Reifung beinhalten, die gametischen Kerne (Pronuklei) erzeugen.
Während der Autogamie in P. aurelia teilen sich die beiden Mikrokerne zweimal (einmal meiotisch), um acht Mikrokerne zu bilden, von denen sechs degenerieren. Währenddessen wächst der Makronukleus zu einer strangförmigen Masse heran, die später in Stücke zerbricht und im Zytoplasma absorbiert wird. Zwei der acht Mikronuklei gehen als Pronuklei in einen protoplasmatischen Kegel ein, der sich in der Nähe des Zellmundes ausbeult.
Die beiden Pronuklei verschmelzen zu einem Synkaryon. Das Synkaryon teilt sich zweimal und bildet vier Mikronuklei. Zwei Mikronuklei werden zu Makronuklei. Das Paramecium und seine Mikrokerne teilen sich und bilden zwei Tochterindividuen mit jeweils einem Makronukleus und zwei Mikrokernen. Dieser Vorgang ist in etwa zwei Tagen abgeschlossen.
Autogamie führt zur Verjüngung der Rasse.
Sie ähnelt der Konjugation insofern, als der neue Makronukleus aus dem Material des Mikronukleus gebildet wird und im neuen Makronukleus die korrekte Chromosomenzahl wiederhergestellt wird; und auch in der Tatsache, dass die Fusion zweier Vorkerne stattfindet. Aber die Autogamie unterscheidet sich von der Konjugation, weil nur ein Individuum an der Autogamie teilnimmt und es beide Vorkerne liefert, es ist eine Art Selbstbefruchtung.
(iii) Hemixis:
Diller (1936) berichtete über die Hemixis bei Paramecium aurelia (Abb. 20.24). Bei der Hemixis handelt es sich in erster Linie um einen Prozess der makrokernigen Fragmentierung und Teilung ohne ungewöhnliche mikronukleare Aktivität. Diller klassifizierte die Hemixis in vier Typen, nämlich A, B, C und D, wie in Abb. 20.24 bei P. aurelia gezeigt, aber er fand auch alle Typen in Massenkulturen von P. caudatum und P. multimicronucleatum.
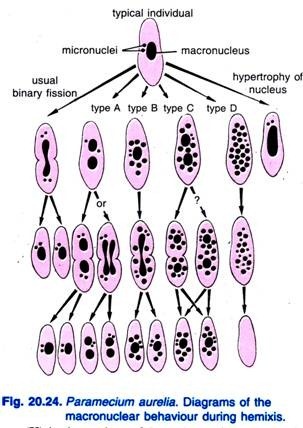
Typ A ist die einfachste Form der Hemixis, die durch eine Teilung des Makronucleus in zwei oder mehr Teile gekennzeichnet ist. Diese Teilung ist nicht mit der Teilung der Mikrokerne synchronisiert.
Typ B ist durch die Extrusion von einem bis 20 oder mehr Chromatinkugeln aus dem Makronukleus in das Zytoplasma gekennzeichnet.
Typ C ist gekennzeichnet durch die gleichzeitige Aufspaltung des Makronukleus in zwei oder mehr Hauptteile und die Extrusion von Makronukleinkugeln in das Zytoplasma.
Typ D gilt als pathologischer Zustand, bei dem der Makronukleus vollständig in Chromatinkügelchen fragmentiert wird, die schließlich aus der Zelle verschwinden. Die Mikrokerne verschwinden im Allgemeinen vor der Auflösung des Makronukleus.
(iv) Zytogamie:
Wichterman (1939) berichtete über einen weiteren sexuellen Prozess in Paramecium caudatum, den er als Zytogamie bezeichnete. Bei der Zytogamie findet kein Kernaustausch statt. Dabei nähern sich zwei Individuen an ihren ventralen Oberflächen an, aber die Pellikel der beiden Individuen bricht nicht ab.
Der Mikronukleus jedes Individuums teilt sich dreimal und bildet acht Mikronuklei, von denen sechs in jedem Individuum zerfallen. Die beiden verbleibenden Mikronuklei verschmelzen zu einem Synkaryon in jeder Zelle. Die Tiere trennen sich nun.
Die Zytogamie unterscheidet sich von der Autogamie dadurch, dass zwei Tiere miteinander in Kontakt stehen, ähnelt aber der Autogamie und der Konjugation durch die Verschmelzung zweier Vorkerne. Die Zytogamie unterscheidet sich von der Konjugation dadurch, dass es keinen Kernaustausch zwischen den beiden zusammenkommenden Tieren gibt.
Einige zytoplasmatische Partikel, die in Paramecium Caudatum gefunden wurden:
(i) Kappa-Partikel:
Im Jahr 1938 berichtete T.M. Sonneborn, dass einige Rassen (bekannt als Killer oder Killerstamm) von Paramecium eine giftige Substanz, genannt Paramecin, produzieren, die für andere Individuen, genannt Sensitive, tödlich ist. Das Paramecin ist wasserlöslich, diffusionsfähig und hängt für seine Produktion von einigen Partikeln ab, die sich im Zytoplasma des Parameciums (Killerstamm) befinden.
Diese Partikel werden Kappa-Partikel genannt. Die Kappa-Partikel enthalten DNA und RNA. Ein Killer-Paramecium kann Hunderte von Kappa-Partikeln enthalten. Die detaillierte Untersuchung dieser Partikel hat ergeben, dass ein dominantes Gen (K) im Kern von Paramecium notwendig ist, damit Kappa-Partikel existieren, sich vermehren und Paramecin produzieren können.
(ii) mµ, Partikel:
R.W. Siegel (1952) berichtete über eine andere Art von Killerpartikeln im Zytoplasma einiger Paramecien. Ein Paramecium mit mp-Partikeln wird als „mate killer“ bezeichnet, weil es, wenn es mit einem Paramecium ohne mµ-Partikel, das als „mate sensitive“ bezeichnet wird, zusammenkommt, letzteres tötet. Die mp-Partikel bestehen auch aus DNA, RNA usw.
Diese Partikel existieren nur in jenen Paramecien, deren Mikronukleus mindestens ein dominantes Gen von einem der beiden Paare von unverbundenen chromosomalen Genen (M1 und M2) enthält.
(iii) Pi-Partikel:
Diese Partikel sollen die mutierte Form der Kappa-Partikel sein, aber sie produzieren keine giftige Substanz.
(iv) Lambda-Teilchen:
Diese Partikel werden in Killer-Paramecien gefunden und sollen eine Substanz produzieren, die für die Lyse oder den Zerfall empfindlicher Paramecien verantwortlich ist, d.h. die diese Substanz nicht besitzen.