- Volvox Case Study Guide Covers up Classification, Characteristics, Volvox Under Microscope, Reproduction (Asexual and Sexual Reproduction)
- Detaillierter Leitfaden mit allen Aspekten
- Volvox Klassifikation
- Volvox Merkmale
- Pflanzenkörper (Der Gametophyt)
- Zellstruktur
- Volvox ein- oder mehrzellig
- Volvox unter dem Mikroskop
- Volvox Fortpflanzung
- Asexuelle Vermehrung
- Sexuelle Fortpflanzung
Volvox Case Study Guide Covers up Classification, Characteristics, Volvox Under Microscope, Reproduction (Asexual and Sexual Reproduction)
Volvox is a fresh water planktonic (free-floating) alga. Es gibt etwa 20 Arten, die zu diesen Gattungen gehören. Im Pflanzenreich erscheint sie als das schönste und attraktivste Objekt. Man findet sie im Süßwasser als grüne, stecknadelkopfgroße Kugeln. Sie sind mit bloßem Auge gerade noch sichtbar und haben einen Durchmesser von etwa 0,5 mm.
Detaillierter Leitfaden mit allen Aspekten
Die Alge wird aufgrund ihres spezifischen Schwimmverhaltens auch als Rollende Alge bezeichnet. Ihr Wachstum wird häufig in temporären oder permanenten Süßwasserteichen, Tümpeln, Gräben und auch in Seen beobachtet. Das Wachstum ist üppig, wenn Temperatur und organische Stoffe in ausreichender Menge vorhanden sind. Seine Vermehrung ist so häufig und reichlich, dass das Wasser der Teiche grün wird (Wasserblüte). Das Frühjahr und die Regenzeit sind die üblichen Zeiten für das aktive vegetative Wachstum. Mit Beginn der ungünstigen Periode (Sommer) verschwindet die Alge und durchläuft eine ungünstige Periode in Form der Zygote. Die Volvox stellen ihre eigene Nahrung durch Photosynthese her.
Volvox Klassifikation
- Klasse: Chlorophyceae
- Ordnung: Volvocales
- Unterordnung: Chlamydomonadineae
- Familie: Sphaerellaceae
- Gattung: Volvox
Die häufigsten Arten von Volvox sind V, globator, V, aureus; V. prolificus, V. rouseletti, V. merelli.
Volvox Merkmale
Pflanzenkörper (Der Gametophyt)
Volvox ist eine coenobiale Grünalge, {(Kolonie-Pflanzenkörper haben keine feste Anzahl von Zellen, z.B. Volvox aureus) (coenobium-Pflanzenkörper mit fester Anzahl von Zellen, z.B., Pandorina moruma, die Anzahl der Zellen beträgt 4, 8, 16 oder 32. Eudorina unicocca, Anzahl der Zellen 16, 32 oder 64)}. Unter den beweglichen Formen sind die Coenobien von Volvox die größten, hoch differenzierten und gut entwickelten Algen. Jedes Coenobium ist ein ellipsoider oder hohler Kugelkörper mit einer genau abgegrenzten, zarten Schleimschicht. Der innere Teil des Coenobiums besteht aus diffluentem (wässrigem) Schleim, während die Zellen am Rand in einer einzigen Schicht angeordnet sind.
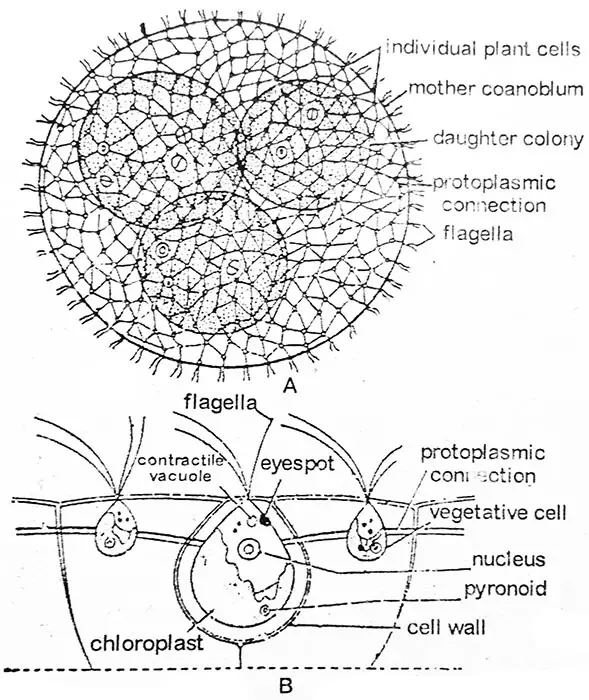
Die Bewegung der Kolonie wird durch die gemeinsame Aktion der Geißeln der einzelnen Zellen bewirkt. Jedes Coenobium hat ein eindeutiges vorderes und ein hinteres Ende. Das Coenobium zeigt Polarität, es bewegt sich und dreht sich langsam, wobei eine bemerkenswerte Zusammenarbeit zwischen den Zellen des vorderen und hinteren Endes im Laufe seiner Bewegung zu beobachten ist. Volvox ist kein Individuum, sondern ein Zusammenschluss einer Reihe ähnlicher Zellen, von denen jede wie ein unabhängiges Individuum funktioniert und ihre eigene Ernährung, Atmung und Ausscheidung durchführt und keine Zusammenarbeit zwischen den Zellen bei diesen Funktionen zeigt. Die Anzahl der Zellen pro Coenobium variiert z.B. 500-1000 bei V. aureus, 1500-20.000 bei V. globator und sogar bis zu ca. 60.000 bei V. rouseletti.
Zellstruktur
Nach der Forschung hat jede einzelne Zelle ihre Identität. Sie ist von einer eigenen, großen, gallertartigen Hülle umgeben, die mit den Hüllen der benachbarten Zellen übereinstimmen oder voneinander verschieden sein kann. Wenn sie getrennt sind, sind sie durch gegenseitige Kompression eckig und haben gewöhnlich einen sechseckigen Umriss. Abb.,2.22. So hilft eine beträchtliche Ausdehnung des gallertartigen Materials, eine Zelle von der anderen zu trennen. Bei den meisten Arten ist jede Zelle mit ihren Nachbarzellen durch eine Reihe von protoplasmatischen oder zytoplasmatischen Strängen (Plasmodesmen, Abb. 2.22) verbunden, die im Laufe der Zellteilungen und der Entwicklung der Kolonie entstehen. Die Protoplasmastränge können bei V. aureus dünn und zart, bei V. globator rund, bei V. mononae keilförmig oder wie bei V. tertius fehlend sein.
Im Umriss ähnelt die einzelne Zelle von Volvox der von Chlamydomopas. Jede Zelle hat anterior ein Paar Geißeln von gleicher Länge. Beide Geißeln sind vom peitschenartigen Typ. Die Geißeln ragen außerhalb der Oberfläche des Coenobiums in das umgebende Wasser. In der Nähe der Basis der Geißeln befinden sich zwei oder mehr kontraktile Vakuolen. Der Protoplast ist je nach Art unterschiedlich geformt. Bei V. tertius ist der Protoplast rundlich und vom Chlamydomonas-Typ, während er bei V. globator sternförmig ist und diffuse Chloroplasten und verstreute kontraktile Vakuolen aufweist.
Volvox ein- oder mehrzellig
Die vegetativen Zellen einer jungen Kolonie sind grün und gleich in Größe und Form, aber in den älteren Kolonien vergrößern sich bestimmte Zellen in der hinteren Region um das Zehnfache oder mehr der Größe der normalen Zellen. Sie entwickeln zahlreiche Pyrenoide, die an Größe zunehmen, und verhalten sich wie Fortpflanzungszellen. Sie können asexuell oder sexuell sein. In einigen Fällen kann ein und dieselbe Kolonie sowohl ungeschlechtliche als auch geschlechtliche Zellen tragen. Im vorderen Teil bleiben die Zellen der Kolonie nur vegetativ und sind vergleichsweise kleiner. In der vorderen Region tragen die Zellen einen größeren Augenfleck. Eine Kolonie besteht also aus zwei Arten von Zellen: Fortpflanzungszellen und somatische Zellen.
Volvox kann als Beispiel für den ersten Schritt zur Koordination und Arbeitsteilung dienen. Eine Volvox-Kolonie kann als mehrzelliger Typus betrachtet werden, der aus Zellen besteht, die für die Ausführung verschiedener Funktionen vorgesehen sind. Die Zellen, die verschiedene Funktionen erfüllen, sind,
- vegetative Zellen, die sich mit der Herstellung von Nahrung befassen und an der Fortbewegung beteiligt sind,
- ungeschlechtliche Zellen, die Tochterkolonien produzieren
- geschlechtliche Zellen: Produktion von Eiern und Produktion von Antheridien
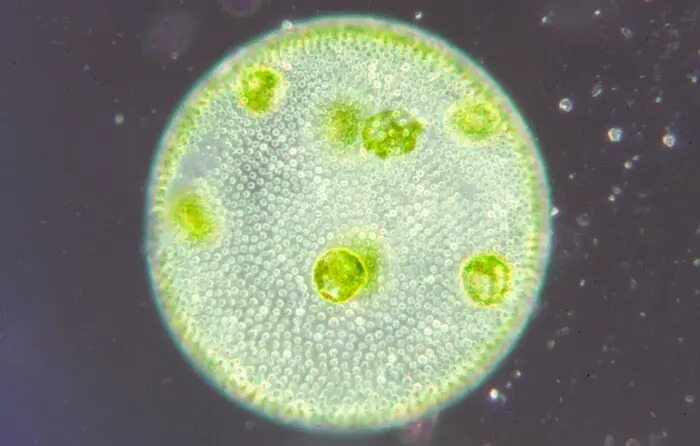
Volvox unter dem Mikroskop
Unten ist die mikroskopische Ansicht einer Kolonie von Volvox:
Volvox Fortpflanzung
Im Gegensatz zu Chlamydomonas zeigen die Zellen der Volvox-Kolonie eine funktionelle Spezialisierung. Sie vermehrt sich sowohl asexuell als auch sexuell. Zu Beginn der Vegetationsperiode (günstige Bedingungen) erfolgt die Vermehrung ungeschlechtlich. Aus diesem Grund sind alle zu einem Zeitpunkt gesammelten Kolonien entweder ungeschlechtlich oder geschlechtlich.
Asexuelle Vermehrung
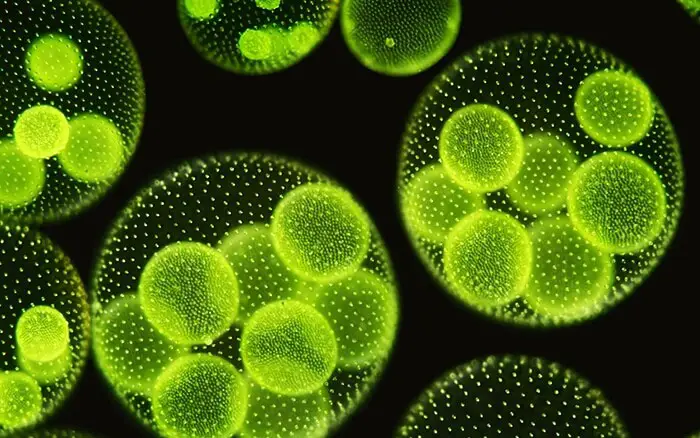
Die ungeschlechtliche Vermehrung findet zu Beginn der Vegetationsperiode unter günstigen Bedingungen statt. In den frühen Stadien sind alle Zellen einer Kolonie gleich, aber später speichern einige Zellen in der hinteren Hälfte der Kolonie die Nahrung und vergrößern sich. Bei diesen stark vergrößerten Zellen handelt es sich um spezialisierte ungeschlechtliche Zellen, die Gonidien (Singular Gonidium) genannt werden. Ihre Zahl schwankt zwischen zwei und fünfzig in einem einzigen Coenobium.
Entwicklung des Tochter-Coenobiums aus dem Gonidium
Vor der Teilung sind die Gonidien leicht in das Innere der Kolonie geschoben und können als eine Reihe vegetativer Zellen durch ihre größere Größe, ihre abgerundete Form, das Fehlen von Geißeln und Augenflecken, einen hervorstehenden Zellkern, mehrere Pyrenoide und ein dichtes körniges Zytoplasma unterschieden werden. Jedes Gonidium liegt in einer kugelförmigen gallertartigen Hülle.
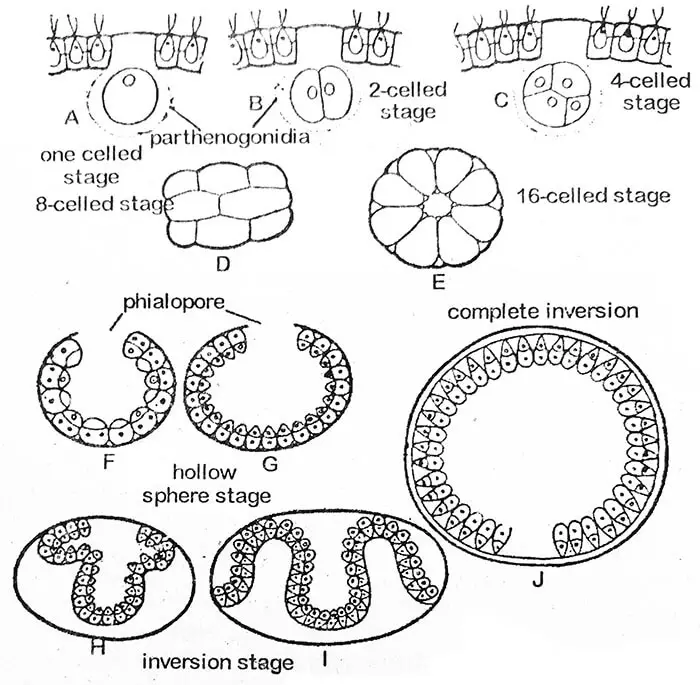
Die erste Teilung des Gonidialprotoplasten ist longitudinal, d.h. in der anterior-posterioren Ebene des Coenobiums. Die zweite Teilung erfolgt ebenfalls in Längsrichtung und im rechten Winkel zur ersten. Jede der so entstandenen Tochterzellen teilt sich erneut in Längsrichtung, so dass eine achtzellige Platte entsteht. Sie wird gekrümmt und ihre konkave Oberfläche zeigt nach außen. Dieses Acht-Zellen-Stadium wird als Plakea-Stadium bezeichnet. Die gleichzeitige Längsteilung der Tochterzellen setzt sich über mehrere Zellgenerationen fort (bis zu 14, 15 oder 16 Mal bei V. rouseletti).
Im Sechzehnzellstadium sind die Zellen innerhalb der Peripherie einer Hohlkugel angeordnet, mit einer kleinen Öffnung, der Phialopore, zur Außenseite des Muttercoenobiums. In diesem Stadium sind alle Zellen nackt und stehen miteinander in Kontakt. Ihre vorderen Enden sind dem Zentrum der Kugel zugewandt. Mit dem Fortschreiten der Invagination vergrößert sich die Phialopore stark. Wenn die Einfaltung des hinteren Teils (Invagination) beginnt, sich durch die Phialopore zu schieben, werden die sie umgebenden Ränder nach hinten gerollt und rutschen allmählich nach unten, bis die gesamte Struktur auf den Kopf gestellt ist. Die Phialopore, die nun eine Reihe von Falten aufweist, wird allmählich geschlossen. Der Prozess der Inversion dauert etwa drei bis fünf Stunden. In einigen abnormalen Fällen findet die Inversion überhaupt nicht statt, wie bei V. minor berichtet wurde.
Die Zellen der Tochterkolonien beginnen nun, sich durch die Entwicklung von schleimigen Teilen (Zellwand) voneinander zu trennen. Jede Zelle erhält schließlich ein Paar Geißeln und eine Zellmembran. Die Tochterkolonie (Coenobium) wird noch von der Mutterzellwand gehalten, die sich schließlich zu einer sie umgebenden schleimigen Membran entwickelt. In einer Mutterkolonie können sich mehrere Tochter-Coenobien gleichzeitig entwickeln. So können sie den hohlen Mittelbereich der Mutterkolonie ausfüllen.
Die Tochter-Coenobien werden mit dem Tod und Zerfall der Mutterkolonie freigesetzt. Manchmal werden die Tochterkolonien über einen längeren Zeitraum nicht freigesetzt und entwickeln Enkelkolonien. So können bis zu 2 bis 4 Generationen von gefangenen Tochterkolonien in einer ursprünglichen Elternkolonie vorkommen, insbesondere bei V. africanus.
Sexuelle Fortpflanzung
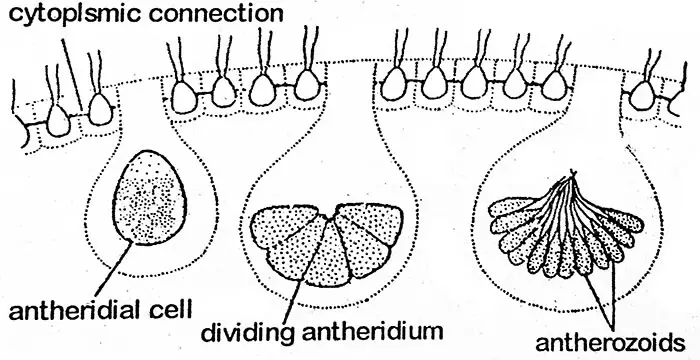
Volvox zeigt eine fortgeschrittene oogame Art der sexuellen Fortpflanzung, die durch die Bildung von Antheridien und Oogonien erfolgt. Sie können auf demselben Coenobium gebildet werden (einhäusig) wie bei V. globator oder auf verschiedenen Coenobien (zweihäusig) wie bei V. aureus. Einhäusige Arten sind protandrisch (die Antheridien entwickeln sich zuerst), so dass bei diesen Arten die Befruchtung zwischen dem Antherozoid und der Eizelle verschiedener Pflanzen stattfindet. Interessant ist, dass sexuelle Kolonien oft keine ungeschlechtlichen Tochterkolonien bilden. In einem Coenobium befinden sich die Zellen, die zur Bildung des Geschlechtsorgans bestimmt sind, in der hinteren Hälfte. Sie werden als spezialisierte Zellen betrachtet. Die Geschlechtsorgane (Gametangien) werden in geringerer Zahl gebildet. Während der Bildung der Gametangien wird die Zelle vergrößert und abgerundet und wirft die Geißeln ab, bleibt aber mit anderen Zellen durch feine Protoplasmafäden verbunden. Das männliche Gametangium wird Antheridium genannt, das weibliche Oogonium.
Der Protoplast eines Antheridiums unterliegt wiederholten Zellteilungen, ähnlich wie bei der Entwicklung einer ungeschlechtlichen Gonidialzelle zu einer Tochterkolonie (d. h. Plakea-Stadium). Auf diese Weise bildet sich eine Masse kleiner, nackter, biflagellater, fusiformer Antherozoiden mit einer Anzahl von 16 bis 512 in einem Antheridium. Diese sind als flache Platte gruppiert, außer bei V. aureus, wo die Antherozoiden in der asexuellen Kolonie zu sehen sind. Die Plakea der Antherozoide löst sich auf und setzt die Antherozoide frei.
Antherozoid
Jedes Antherozoid ist eine biflagellate, längliche, konische oder fusiforme Struktur mit einem einzigen Kern und einem kleinen gelbgrünen oder blassgrünen Chloroplasten.
Oogonium
Die Oogoniumzelle vergrößert sich beträchtlich und wirft ihre Geißeln und protoplasmatischen Verbindungen mit den Nachbarzellen ab. Die Zelle wird rundlich oder kolbenförmig und ragt mit einem Großteil ihres Teils in das Innere des Coenobiums hinein. In diesem Stadium wird sie als Oogonium bezeichnet, dessen gesamter Teil sich in ein einziges kugelförmiges Ei mit einem schnabelartigen Vorsprung an einer Seite verwandelt. Durch dieses Ende dringt das Antherozoid in das Oogonium ein. Das Ei (Oosphäre) enthält einen großen, zentral gelegenen Zellkern und einen parietalen Chloroplasten mit Pyrenoiden. Es ist reichlich mit Reservestoffen versehen, die oft von den benachbarten Zellen durch protoplasmatische Stränge aufgenommen werden.
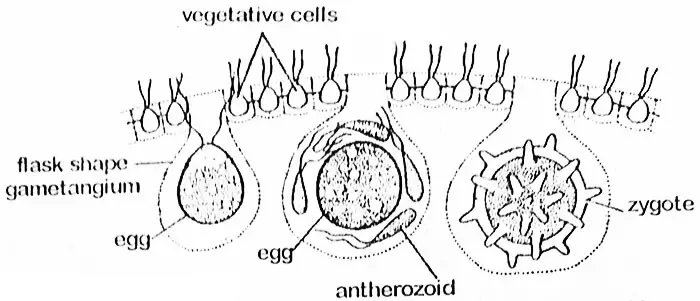
Befruchtung
Die Antherozoide werden zum Zeitpunkt der Befruchtung in Gruppen freigesetzt und bleiben intakt, bis sie das Ei erreichen. Dann werden die Antherozoide freigesetzt. Nur ein Antherozoid verschmilzt mit dem Ei und führt zur Bildung einer Oospore. Die Oospore sondert anschließend eine dreischichtige glatte oder stachelige Wand ab. Sie akkumuliert genügend Hämatochrom (rote Farbpigmentkörnchen, die wahrscheinlich aus Xanthophyll bestehen), das ihr ein orangefarbenes Aussehen verleiht. In diesem Stadium kann sie als Zygote bezeichnet werden.
Oospore und ihre Keimung
Die äußere Wand, Exospore, kann glatt (V. globator) oder stachelig (V. speematospaera) sein. Die mittlere Schicht ist die Mesospore und die innere die Endospore. Die Zygote enthält genügend Reserven an Nahrungsmaterial und anderen Einschlüssen. So wird die Zygote vom Coenobium zurückgehalten, das durch den Zerfall der gallertartigen Matrix freigesetzt werden kann. Nach der Freisetzung setzt sie sich am Boden des Beckens ab und kann mehrere Jahre lang lebensfähig bleiben.

Bei Eintritt günstiger Bedingungen entwickelt sich die Zygote auf unterschiedliche Weise. Bei V. campensis teilt sich der Zygotenkern meiotisch und bildet vier Kerne, von denen drei degenerieren und einer überlebt: Der überlebende Kern entweicht zusammen mit dem Zytoplasma aus der Blase. In diesem Stadium kann er als Schwärmer (große Anzahl oder dichte Gruppe von Insekten, Zellen usw.) bezeichnet werden. Er schwimmt frei und teilt sich und teilt sich wieder, um ein neues Coenobium zu bilden. Während der Keimung werden die beiden äußeren Wandschichten gallertartig und die innere Schicht bildet ein Vesikel, das später mit dem Protoplasten der Zygote gefüllt wird. Bei V. rouseletti und V. minor wird der Protoplast der Zygote in eine einzelne Zoospore umgewandelt, die durch weitere Teilungen ein neues Coenobium bildet. Ein solches Coenobium besteht aus einer kleineren Anzahl von Zellen, die sich in den nächsten sechs oder mehr Generationen ungeschlechtlich fortpflanzen, wobei sich die Anzahl in den folgenden Generationen jedes Mal erhöht.
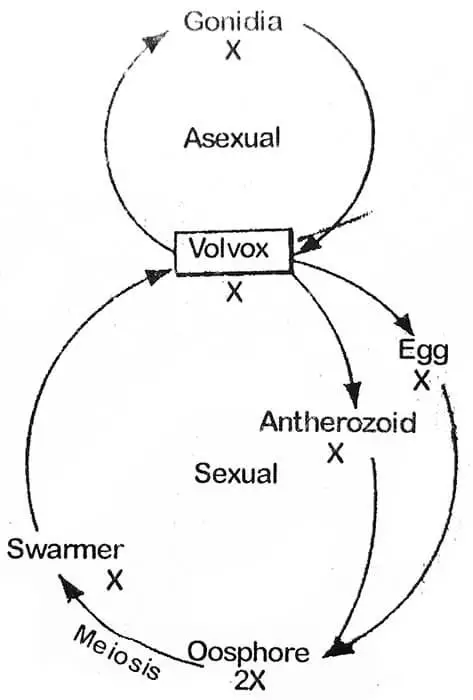
Die Zygote ist die einzige diploide Phase im Lebenszyklus von Volvox und daher ist der Hauptkörper der Pflanze haploid. Aus diesem Grund muss die Zygote bei der Bildung einer neuen Kolonie eine Reduktionsteilung durchlaufen.