Vue d’ensemble des gamètes et de la fécondation
La reproduction sexuée désigne les cellules germinales sexuelles (gamètes) produites par les parents, par la combinaison de cellules germinales bisexuées (telles que les spermatozoïdes et les ovules), deviennent l’ovule fécondé, puis l’ovule fécondé se développe en un nouvel individu, appelé reproduction sexuée. Une variation importante des combinaisons de gènes dans la reproduction sexuée peut augmenter la capacité de la progéniture à s’adapter à la sélection naturelle. Les gènes combinés de manière aléatoire dans la progéniture de la reproduction sexuelle peuvent ou non être bénéfiques pour l’espèce, mais ils augmentent au moins les chances de survie de quelques individus dans un environnement imprévisible et en constante évolution, ce qui profite à l’espèce. La reproduction sexuée peut également favoriser la propagation de mutations favorables dans la population. Si deux individus d’une espèce possèdent des mutations favorables à des endroits différents, dans la population à reproduction asexuée, les deux mutants se feront concurrence jusqu’à une élimination, il est impossible de conserver les deux mutations favorables en même temps. Cependant, dans les populations à reproduction sexuée, grâce à l’accouplement et à la recombinaison, ces deux mutations favorables peuvent entrer simultanément dans le génome d’un même individu et se répandre simultanément dans la population. Pour les raisons susmentionnées, la reproduction sexuée accélère le processus d’évolution. Au cours des plus de 3 milliards d’années d’évolution biologique sur Terre, les deux premiers milliards d’années de vie sont restés au stade de la reproduction asexuée, et l’évolution a été lente, et le taux d’évolution s’est considérablement accéléré depuis le dernier milliard d’années. Outre les changements dans l’environnement terrestre (tels que l’émergence de l’atmosphère contenant de l’oxygène, etc.), l’apparition et le développement de la reproduction sexuée en sont également une cause majeure.
Les gamètes
Les gamètes désignent les cellules matures produites par le système reproducteur lorsque l’organisme se reproduit sexuellement, appelées cellules germinales. Les gamètes se divisent en gamètes mâles et gamètes femelles. Les gamètes femelles des animaux et des plantes sont généralement appelés ovules, et les gamètes mâles sont appelés spermatozoïdes. Le sperme est assez petit, mais il peut se déplacer et entrer dans l’ovule en forme de scorpion. La cellule œuf est assez grande et ne nage pas. Par exemple, le volume de l’ovule de l’oursin est 10 000 fois supérieur à celui des spermatozoïdes. Bien que les gamètes mâles et femelles soient de volume différent, l’ADN nucléaire qu’ils fournissent à la progéniture est égal, c’est-à-dire que chacun fournit un ensemble de génomes. Cependant, en raison de la grande taille des ovules, la structure cytoplasmique et l’ADN cytoplasmique des cellules filles sont essentiellement fournis par les ovules. Les gamètes jouent un rôle important dans les calculs biologiques. Grâce aux cartes génétiques, le flux des gènes et le modèle des génotypes de la descendance peuvent être clairement observés. Le nombre de gamètes dans la première génération d’hybrides et les traits relatifs dans les hybrides sont de puissance 2nème (n est le logarithme des traits relatifs). Lorsqu’un organisme vivant forme un gamète, les facteurs génétiques appariés sont séparés les uns des autres et entrent dans des gamètes différents. Les gamètes ne contiennent qu’un seul de chaque paire de facteurs génétiques. Lors de la fécondation, la combinaison des gamètes mâles et femelles est aléatoire. En raison du genre de gamètes formés par la méiose, de la diversité de la composition des chromosomes, de la différence de matériel génétique des différents gamètes et du caractère aléatoire de la combinaison des ovules et des spermatozoïdes dans le processus de fécondation, la progéniture d’un même parent doit être diverse.
Fécondation
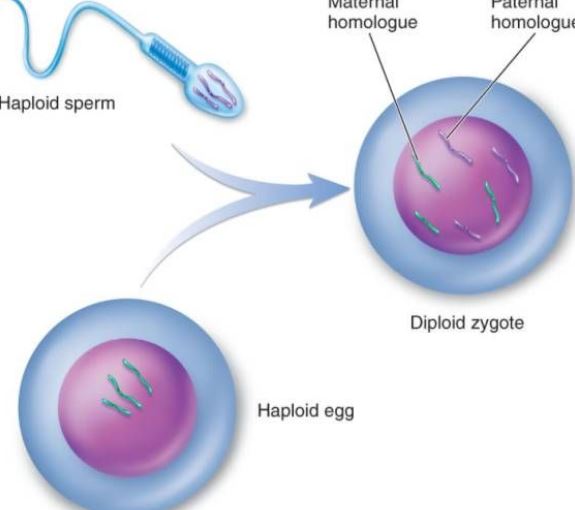
La fécondation est le processus par lequel les ovules et les spermatozoïdes fusionnent en un zygote. C’est une caractéristique de base de la reproduction sexuée et elle est omniprésente dans le monde animal et végétal, mais on parle souvent de l’animal. Fécondation animale Au niveau cellulaire, le processus de fécondation comprend trois grandes étapes : l’activation de l’ovule, sa régulation et la fusion pronucléaire amphotère. L’activation peut être considérée comme le point de départ de l’ontogenèse, qui se manifeste principalement par des changements dans la perméabilité de la membrane suintante, l’ecclésia des granules corticaux, la formation de la membrane fécondée, etc. L’ajustement se produit après l’activation, qui est la première étape pour assurer la division normale des œufs fécondés. Le changement du noyau procaryote assure l’héritage des parents et rétablit le diploïde. La fécondation ne déclenche pas seulement la réplication de l’ADN mais active également les informations génétiques telles que l’ARNm et l’ARNr dans l’ovule pour synthétiser les protéines nécessaires au développement de l’embryon. Six à sept jours après la fécondation, la zone pellucide du blastocyste disparaît et s’enfouit progressivement pour être recouverte par l’endomètre, ce qui s’appelle l’implantation de l’ovule fécondé. Le sperme d’un animal n’a pas de chimiotaxie évidente comme le sperme des plantes inférieures telles que les bryophytes, mais atteint le voisinage de l’ovule par un mouvement actif ou par le mouvement ciliaire des cellules épithéliales génitales. La capacitation des spermatozoïdes : On sait que lorsque de nombreux spermatozoïdes de mammifères traversent l’appareil reproducteur féminin ou le cumulus, la protéine étrangère qui encapsule le sperme est éliminée, et les propriétés physiques et biologiques de la membrane plasmique du sperme changent, ce qui permet au sperme de gagner de l’énergie et de participer au processus de fécondation. Lorsque le spermatozoïde capacitif de mammifère entre en contact avec la membrane de l’ovule ou la zone pellucide de l’ovule, il se lie spécifiquement à une glycoprotéine sur la membrane de l’ovule, ce qui stimule la production d’énergie par le spermatozoïde. La réaction acrosomique aide le spermatozoïde à traverser davantage la membrane de l’ovule. Sur l’œuf d’oursin, la réaction acrosomique du spermatozoïde est due à une certaine substance polysaccharide dans le film péri-membranaire. La plupart des œufs ont des membranes d’œuf sur la périphérie, et l’épaisseur des différentes membranes d’œuf varie. Le composant principal est la mucine ou le mucopolysaccharide ; seuls quelques uns sont des œufs nus, comme les œufs de coelentérés. Réaction de l’acrosome : Lorsque le spermatozoïde traverse la membrane de l’ovule, il y a un processus de liaison. Le premier est faiblement attaché, sans interférence de la température externe, et n’a pas de spécificité. Pendant la période d’adhésion, la protéine du protoplaste sur la membrane acrosomique se transforme en protéine acrosomique, et la protéine acrosomique accélère la traversée de la membrane de l’ovule par les spermatozoïdes ; il s’agit d’une combinaison forte, qui peut être interférée par une température basse, et qui a une spécificité. Une protéine qui se lie spécifiquement à la glycoprotéine de l’ovule a été isolée sur la membrane plasmique du sperme de l’oursin, appelée protéine de liaison, avec un poids moléculaire d’environ 30 000. Les ovocytes secondaires au milieu de la deuxième division de maturation pendant l’ovulation, ainsi que la zone pellucide environnante et la couronne de radiation, pénètrent rapidement dans l’ampoule de la trompe de Fallope à travers la cavité abdominale grâce au balancement des cils des cellules épithéliales de la trompe de Fallope et à la contraction de la couche musculaire. Si l’ovule ne rencontre pas le spermatozoïde, il commence généralement à dégénérer dans les 12 à 24 heures. Une fois que le spermatozoïde est en contact avec l’ovule, l’ovule lui-même subit une série de changements d’activation. Sur les œufs de mammifères, elle se caractérise par la réaction corticale, la réaction de la membrane ovarienne et la réaction de la zone pellucide, qui peuvent bloquer la multifécondation et stimuler le développement ultérieur des œufs. La réaction corticale se produit au moment de la fusion des spermatozoïdes. À partir du point de fusion, les particules corticales se rompent et le contenu est efflué, étalant ainsi le cortex de l’ovule entier. La réaction de la membrane ovarienne est un processus de recombinaison de l’enveloppe de l’ovule et des granules corticaux. La réaction de la zone pellucide est un processus dans lequel l’efflux cortical et la zone pellucide d’une membrane fécondée, la membrane de l’ovule est séparée de la membrane plasmique, le récepteur des spermatozoïdes dans la zone pellucide disparaît, et la zone pellucide durcit. Seuls les spermatozoïdes qui ont une réaction acrosomique peuvent fusionner avec l’ovule. Sous l’action de l’acrosine, le spermatozoïde traverse la couronne radiale et interagit avec la molécule glycoprotéique ZP3 du récepteur du sperme sur la zone pellucide, ce qui permet au spermatozoïde de libérer l’enzyme acrosomique et de pénétrer dans la périvitelline à travers la zone pellucide. Au début de la fécondation, l’enveloppe de la partie équatoriale de la tête du spermatozoïde humain est en contact avec la membrane de l’ovule, puis le noyau et le cytoplasme du spermatozoïde entrent dans l’ovule. Après l’entrée du spermatozoïde dans l’ovule, les particules corticales dans le cytoplasme peu profond de l’ovule libèrent immédiatement son contenu à la périphérie de la membrane. Dans le même temps, lorsque la membrane des granules corticaux fusionne avec les ovules, la charge superficielle négative de la surface de la cellule augmente, supprimant ainsi la fusion de la membrane plasmique du spermatozoïde avec la membrane de l’ovule, ce que l’on appelle une réaction corticale. La structure de la zone pellucide se modifie, ce que l’on appelle la réaction de la zone pellucide. À ce moment, la capacité de la zone pellucide à fixer les spermatozoïdes est réduite, ce qui empêche l’apparition de la polyspermie et garantit les caractéristiques biologiques de la monospermie humaine. En plus de détruire ou d’inactiver le récepteur lié aux spermatozoïdes dans la zone pellucide, le contenu des granules corticaux peut également augmenter la réticulation entre les chaînes peptidiques dans la zone pellucide, affaiblir la sensibilité à la protéase de l’acrosome et empêcher la pénétration des spermatozoïdes. Bien que plusieurs spermatozoïdes traversent la zone pellucide, un seul d’entre eux pénètre dans l’ovule pour le féconder. Dans une situation anormale, deux spermatozoïdes participent à la fécondation, c’est la fécondation par double sperme. Deux spermatozoïdes entrent simultanément dans l’ovule pour former des embryons de cellules triploïdes, qui avortent tous ou meurent peu après la naissance. Après l’entrée du spermatozoïde dans l’ovule, celui-ci accomplit rapidement la deuxième division mature. À ce moment, les noyaux du spermatozoïde et de l’ovule sont appelés respectivement pronucléus mâle et pronucléus femelle. Les deux pronucléus se rapprochent progressivement, la membrane nucléaire disparaît et les chromosomes fusionnent pour former un ovule fécondé diploïde. Lorsque les spermatozoïdes sont fusionnés, on peut voir les microvillosités à la surface de l’ovule entourer le spermatozoïde, qui peut être orienté ; puis la membrane ovarienne fusionne avec la membrane plasmique dans la région postérieure de l’acrosome du spermatozoïde.
Référence
- Yanagimachi R. Germ cells and fertilization : why I studied these topics and what I learned along the path of my study. Andrologie. 2014, 2(6):787-93.
- Henshaw J M, Marshall D J, Jennions M D, et al. La compétition locale des gamètes explique l’allocation des sexes et les stratégies de fertilisation dans la mer. Naturaliste américain. 2014, 184(2):32.
- Mori T, Igawa T. Processus de fixation des gamètes révélé dans la fertilisation des plantes à fleurs. Signalisation des plantes &Comportement. 2014, 9(12):977715.
- Hédouin L, Pilon R, Puisay A. Le stress d’hyposalinité compromet la fécondation des gamètes plus que la survie des larves de corail. Recherche sur l’environnement marin. 2015, 104:1-9.
Luo J, Mcginnis L K, Carlton C, et al. Fonction de PTK2b pendant la fécondation de l’ovocyte de souris. Biochem Biophys Res Commun. 2014, 450(3):1212-1217.