- Le guide d’étude de cas Volvox couvre up Classification, caractéristiques, Volvox au microscope, reproduction (reproduction asexuée et sexuelle)
- Guide détaillé avec tous les aspects
- Volvox Classification
- Caractéristiques de Volvox
- Corps végétal (le gamétophyte)
- Structure cellulaire
- Volvox Unicellulaire ou Multicellulaire
- Volvox au microscope
- Reproduction du Volvox
- Reproduction asexuée
- Reproduction sexuée
Le guide d’étude de cas Volvox couvre up Classification, caractéristiques, Volvox au microscope, reproduction (reproduction asexuée et sexuelle)
Volvox est une algue planctonique (flottant librement) d’eau douce. Il existe environ 20 espèces appartenant à ces genres. Dans le règne végétal, elle apparaît comme l’objet le plus beau et le plus attrayant. On la trouve dans l’eau douce sous forme de boules vertes de la taille d’une tête d’épingle. Elles sont juste visibles à l’œil nu, d’environ 0,5 mm de diamètre.
Guide détaillé avec tous les aspects
L’algue en raison de son mode de nage spécifique est souvent appelée, algue roulante. Sa croissance est fréquemment observée dans les étangs d’eau douce temporaires ou permanents, les piscines, les fossés et également dans les lacs. La croissance est abondante lorsque la température et la matière organique sont disponibles en quantité suffisante. Sa multiplication est si fréquente et abondante que l’eau des étangs devient verte (water bloom). Le printemps et la saison des pluies sont les périodes habituelles de sa croissance végétative active. A l’arrivée d’une période défavorable (été) l’algue disparaît et passe une période défavorable sous forme de zygote. Les volvox fabriquent leur propre nourriture par photosynthèse.
Volvox Classification
- Classe : Chlorophycées
- Ordre : Volvocales
- Sous-ordre : Chlamydomonadineae
- Famille : Sphaerellaceae
- Genre : Volvox
Les espèces les plus courantes de Volvox sont V, globator, V, aureus ; V. prolificus, V. rouseletti, V. merelli.
Caractéristiques de Volvox
Corps végétal (le gamétophyte)
Volvox est une algue verte cénobiale, {(colonie-corps végétal n’ont pas un nombre fixe de cellules, par exemple Volvox aureus) (cénobium-corps végétal ayant un nombre fixe de cellules, par exemple, Pandorina moruma, le nombre de cellules est de 4, 8, 16 ou 32. Eudorina unicocca, nombre de cellules 16, 32 ou 64)}. Parmi les formes mobiles, les coenobiums de Volvox sont les algues les plus grandes, les plus différenciées et les plus évoluées. Chaque coenobium est un corps ellipsoïde ou une sphère creuse avec une couche définie de mucilage délicat exactement marquée. La partie intérieure du coenobium est composée de mucilage diffluent (aqueux), tandis que les cellules sont disposées en une seule couche à la périphérie.
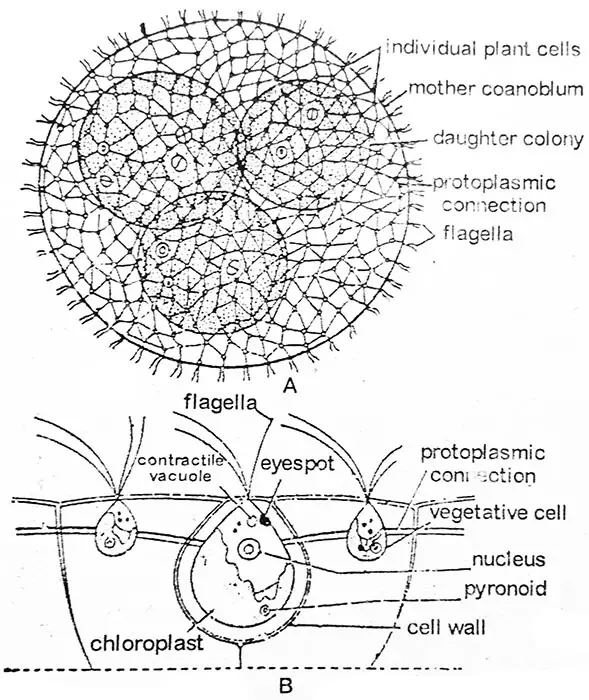
Le mouvement de la colonie est provoqué par l’action conjointe des flagelles des cellules individuelles. Chaque coenobium a une extrémité antérieure et une extrémité postérieure bien définies. Le coenobium présente une polarité, il se déplace et tourne lentement, montrant une coopération remarquable entre les cellules des extrémités antérieure et postérieure au cours de son mouvement. Le Volvox n’est pas un individu mais une association d’un certain nombre de cellules similaires, dont chacune fonctionne comme un individu indépendant et effectue sa propre nutrition, respiration et excrétion et ne montre aucune coopération entre les cellules dans ces fonctions. Le nombre de cellules par coenobium varie par exemple de 500 à 1000 chez V. aureus, 1500 à 20 000 chez V. globator et même jusqu’à environ 60 000 chez V. rouseletti.
Structure cellulaire
Selon les recherches, chaque cellule individuelle a son identité. Elle est entourée de sa propre grande gaine, gélatineuse, qui peut être en conflit avec les gaines des cellules adjacentes ou être distincte les unes des autres. Lorsqu’elles sont distinctes, elles sont anguleuses par compression mutuelle et ont généralement un contour hexagonal. Fig. 2.22. Ainsi, une étendue considérable de matière gélatineuse permet de séparer une cellule d’une autre. Dans la majorité des espèces, chaque cellule est reliée à ses voisines par une série de brins protoplasmiques ou cytoplasmiques (plasmodesmae Fig. 2.22) établis au cours des divisions cellulaires et du développement de la colonie. Les brins protoplasmiques peuvent être fins et délicats chez V. aureus, ronds chez V. globator ; ou cunéiformes chez V. mononae, ou peuvent être absents comme chez V. tertius.
Dans son contour, la cellule individuelle de Volvox ressemble à Chlamydomopas. Chaque cellule a inséré antérieurement une paire de flagelles de longueur égale. Les deux flagelles sont de type fouetté. Les flagelles se projettent à l’extérieur de la surface du coenobium dans l’eau environnante. Près de la base des flagelles, deux vacuoles contractiles ou plus sont présentes. Le protoplaste a des formes différentes selon l’espèce. Chez V. tertius le protoplaste est arrondi et de type Chlamydomonas, alors que chez V. globator le protoplaste est de type stellaire ayant un chloroplaste diffus et des vacuoles contractiles dispersées.
Volvox Unicellulaire ou Multicellulaire
Les cellules végétatives d’une jeune colonie sont vertes et semblables en taille et en forme mais dans les colonies plus anciennes, certaines cellules de la région postérieure augmentent dix fois ; ou plus la taille de la cellule normale. Elles développent de nombreux pyrénoïdes dont la taille augmente et se comportent comme des cellules reproductrices. Elles peuvent être asexuées ou sexuées. Dans certains cas, la même colonie peut porter à la fois des cellules asexuées et sexuées. Dans la partie antérieure, les cellules de la colonie restent uniquement végétatives et sont comparativement plus petites. Dans la région antérieure, les cellules portent une tache oculaire plus grande. Ainsi, une colonie est constituée de deux types de cellules : les cellules reproductrices et les cellules somatiques.
Volvox peut servir d’exemple pour le premier pas vers la coordination et la division du travail. Une colonie de Volvox peut être considérée comme un type multicellulaire composé de cellules mises à part pour l’exécution de différentes fonctions. Les cellules remplissant différentes fonctions sont,
- Cellules végétatives concernées par la fabrication de la nourriture et impliquées dans la locomotion,
- Cellules asexuées produisant des colonies filles
- Cellules sexuées : produisant des œufs, et produisant des anthéridies
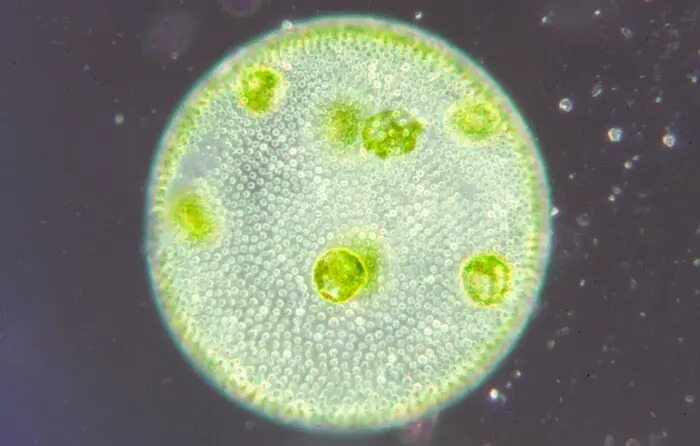
Volvox au microscope
Voici la vue microscopique d’une colonie de volvox :
Reproduction du Volvox
Contrairement à Chlamydomonas, les cellules de la colonie de Volvox montrent une spécialisation fonctionnelle. Elle se reproduit à la fois de manière asexuée et sexuée. Au début de la saison de croissance (conditions favorables). la reproduction est asexuée. C’est pour cette raison que toutes les colonies collectées à un moment donné sont soit asexuées, soit sexuées.
Reproduction asexuée
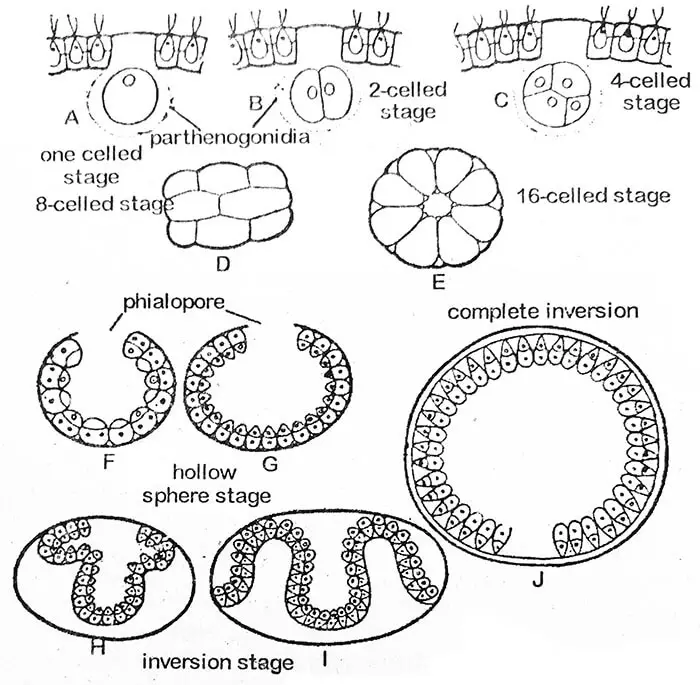
La reproduction asexuée a lieu au début de la saison de croissance pendant les conditions favorables. Dans les premiers stades, toutes les cellules d’une colonie sont semblables mais, plus tard, quelques cellules de la moitié postérieure de la colonie stockent la nourriture et augmentent en taille. Ces cellules fortement agrandies sont des cellules asexuées spécialisées appelées gonidies (singulier gonidium). Leur nombre varie de deux à cinquante dans un seul coenobium.
Développement du coenobium fille à partir du gonidium
Avant la division, les gonidia sont légèrement poussés à l’intérieur de la colonie et peuvent être distingués comme une rangée de cellules végétatives par leur plus grande taille, leur forme arrondie, l’absence de flagelles et de tache oculaire, un noyau proéminent, plusieurs pyrénoïdes et un cytoplasme granulaire dense. Chaque gonidium se trouve dans une gaine gélatineuse globulaire.
La première division du protoplaste gonidial est longitudinale c’est-à-dire dans le plan antéro-postérieur du coenobium. La deuxième division est également longitudinale et à angle droit par rapport à la première. Chacune des cellules filles ainsi formées se divise à nouveau dans le sens de la longueur, de sorte qu’une plaque de huit cellules est formée. Elle devient incurvée avec sa surface concave tournée vers l’extérieur. Ce stade à huit cellules est connu sous le nom de stade Plakea. Les divisions longitudinales simultanées des cellules filles se poursuivent pendant plusieurs générations cellulaires (jusqu’à 14, 15 ou 16 fois chez V. rouseletti).
Au stade de seize cellules, les cellules sont disposées à la périphérie d’une sphère creuse, avec une petite ouverture, le phialopore vers l’extérieur du coenobium parent. A ce stade, toutes les cellules sont nues et en contact les unes avec les autres. Leurs extrémités antérieures sont tournées vers le centre de la sphère. Avec la progression de l’invagination, le phialopore s’agrandit considérablement. Lorsque le pliage intérieur de la partie postérieure (invagination) commence à pousser à travers le phialopore, les bords qui l’entourent se recourbent vers l’arrière et glissent progressivement vers le bas jusqu’à ce que la structure entière soit inversée. Le phialopore, qui présente maintenant un certain nombre de plis, se ferme progressivement. Le processus d’inversion prend environ trois à cinq heures. Dans certains cas anormaux, l’inversion n’a pas lieu du tout, comme cela a été signalé chez V. minor.
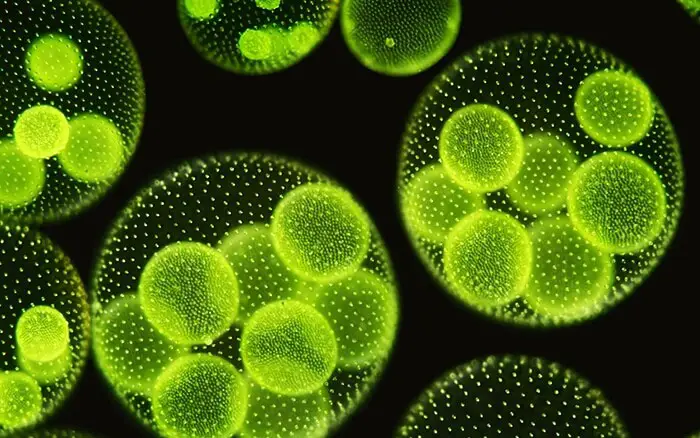
Les cellules des coenobiums filles commencent maintenant à se séparer les unes des autres par le développement de portions mucilagineuses (paroi cellulaire). Chaque cellule, enfin, acquiert une paire de flagelles et une membrane cellulaire. La colonie fille (coenobium) est toujours retenue dans la paroi cellulaire parentale qui finit par se transformer en une membrane mucilagineuse qui l’entoure. Plusieurs coenobiums filles peuvent se développer simultanément dans une colonie mère. Ainsi, ils peuvent remplir la région centrale creuse de la colonie mère.
Les coenobiums filles sont libérés avec la mort et la décomposition de la colonie mère. Parfois, les colonies filles ne sont pas libérées pendant une plus longue durée et développent des colonies petites-filles. Ainsi, jusqu’à 2 à 4 générations de colonies filles emprisonnées peuvent être observées dans une colonie parentale originale, en particulier chez V. africanus.
Reproduction sexuée
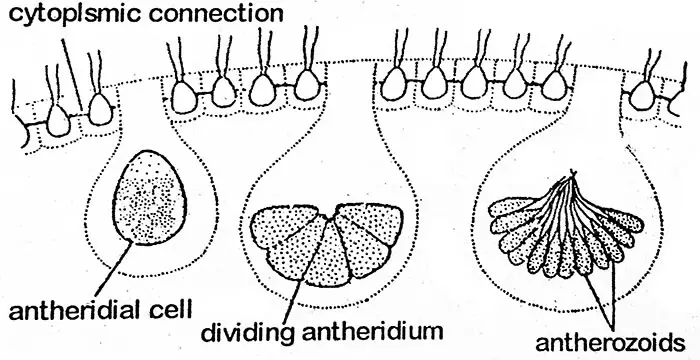
Volvox présente une reproduction sexuée avancée de type oogame qui a lieu par la formation d’anthéridies et d’oogonies. Ils peuvent être formés sur le même coenobium (monoïque) comme chez V. globator ou sur des coenobiums différents (dioïque) comme chez V. aureus. Les espèces monoïques sont protandres (les anthéridies se développent en premier) donc, dans ces espèces, la fertilisation se fera entre l’anthérozoïde et l’ovule de plantes différentes. Il est assez intéressant de constater que les colonies sexuées sont souvent dépourvues de colonies filles formées asexuellement. Chez un coenobium, les cellules destinées à former l’organe sexuel sont présentes dans la moitié postérieure. Elles sont considérées comme des cellules spécialisées. Les organes sexuels (gamétanges) sont produits en nombre réduit. Pendant la formation des gamétanges, la cellule s’élargit et s’arrondit et se débarrasse de ses flagelles, mais elle reste reliée aux autres cellules par de fins fils protoplasmiques. Le gamétangue mâle est appelé anthéridie tandis que l’oogonium femelle.
Le protoplaste d’une anthéridie subit des divisions cellulaires répétées d’une manière similaire à celle observée dans le développement d’une cellule gonidiale asexuée en une colonie fille (c’est-à-dire le stade plakea). Ainsi, une masse. de petits anthérozoïdes fusiformes, nus, biflagellés, au nombre de 16 à 512 dans un anthéridie est formée. Ils sont regroupés en une plaque plate, sauf chez V. aureus où les anthérozoïdes sont observés dans une colonie asexuée. Le plakea des anthérozoïdes se dissocie et libère les anthérozoïdes.
Antherozoïde
Chaque anthérozoïde est une structure biflagellée, allongée, conique ou fusiforme avec un seul noyau et un petit chloroplaste jaune-vert ou vert pâle.
Oogonium
La cellule oogoniale s’agrandit considérablement et se défait de ses flagelles et de ses connexions protoplasmiques avec les cellules voisines. La cellule devient arrondie ou en forme de flasque avec une grande partie de sa portion faisant saillie à l’intérieur du coenobium. À ce stade, on parle d’oogonium dont toute la partie est transformée en un seul œuf sphérique avec une protubérance en forme de bec vers un côté. Par cette extrémité, l’anthérozoïde pénètre dans l’oogonium. L’œuf (oosphère) contient un gros noyau placé au centre et un chloroplaste pariétal avec des pyrénoïdes. Il est abondamment pourvu de substances de réserve souvent absorbées des cellules voisines par des brins protoplasmiques.
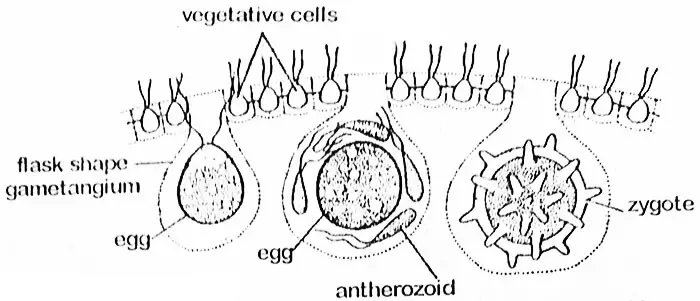
Fertilisation
Les anthérozoïdes sont libérés en groupes au moment de la fertilisation et ceux-ci restent intacts jusqu’à ce qu’ils atteignent l’œuf. Les anthérozoïdes sont alors libérés. Un seul anthérozoïde fusionne avec l’ovule et entraîne la formation d’une oospore. L’oospore sécrète ensuite une paroi à trois couches, lisse ou épineuse. Elle accumule suffisamment d’hématochrome (granules de pigment de couleur rouge probablement de nature xanthophylle) qui lui donne une apparence orange. A ce stade, il peut être appelé zygote.
Oospore et sa germination
La paroi externe, l’exospore peut être lisse, (V. globator) ou épineuse (V. speematospaera). La couche intermédiaire est la mésospore et l’interne est l’endospore. Le zygote contient une réserve suffisante de matière alimentaire et d’autres inclusions. Ainsi, le zygote est retenu par le coenobium qui peut être libéré par la désintégration de la matrice gélatineuse. Après sa libération, il se dépose au fond du bassin et peut rester viable pendant plusieurs années.

A l’apparition de conditions favorables, le zygote se développe de différentes manières. Chez V. campensis le noyau du zygote se divise méiotiquement et forme quatre noyaux, trois d’entre eux dégénèrent et un survit : Le noyau survivant accompagné du contenu cytoplasmique s’échappe de la vésicule. A ce stade, il peut être désigné comme swarmer (grand nombre ou groupe dense, d’insectes, de cellules etc.) il nage librement et se divise et se re-divise pour former un nouveau coenobium. Pendant la germination, les deux couches extérieures de la paroi deviennent gélatineuses et la couche intérieure forme une vésicule qui se remplit ensuite du protoplaste du zygote. Chez V. rouseletti et V. minor, le protoplaste du zygote est converti en une seule zoospore qui, par d’autres divisions, forme un nouveau coenobium. Un tel coenobium se compose d’un plus petit nombre de cellules qui se reproduit asexuellement pour les six générations suivantes ou plus, augmentant chaque fois le nombre dans les générations suivantes.
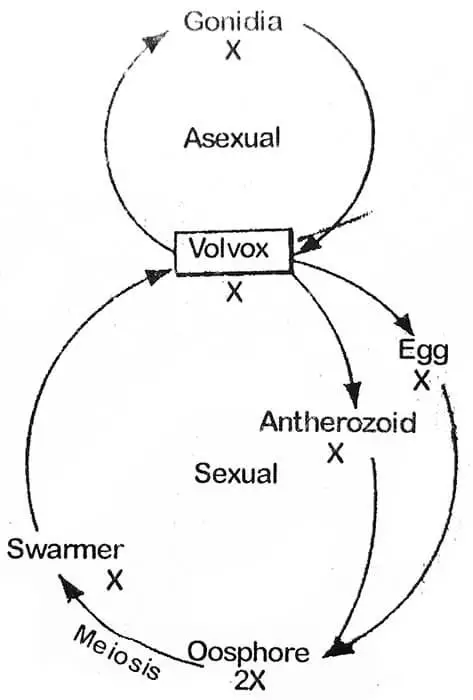
Le zygote est la seule phase diploïde du cycle de vie du Volvox et, par conséquent, le corps principal de la plante est haploïde. C’est pourquoi le zygote doit subir une division réductrice lors de la formation d’une nouvelle colonie.