La dopamine est un produit chimique organique de la famille des catécholamines et des phénéthylamines. La dopamine fonctionne comme un neurotransmetteur dans le cerveau.
La signalisation dopaminergique est associée au comportement motivé par la récompense et au contrôle moteur, le dysfonctionnement du système dopaminergique entraînant de nombreuses maladies. Par exemple, la maladie dégénérative de Parkinson est causée par la perte de neurones sécrétant de la dopamine, ce qui entraîne une déficience motrice. Enzo Life Sciences propose un kit ELISA Dopamine pour la mesure quantitative de la concentration de dopamine dans le sérum, le plasma et les surnageants de culture cellulaire.
Découverte initiale du rôle de la dopamine en tant que neurotransmetteur
Avant 1957, le point de vue dominant était que la 3-hydroxytyramine était un intermédiaire dans la synthèse de la noradrénaline et de l’adrénaline à partir de la tyrosine. Cependant, entre 1957 et 1959, les efforts parallèles de Kathleen Montagu et de ses collègues du laboratoire de Hans Weil-Malherbe à l’hôpital Runwell (Angleterre) et d’Arvin Carlsson et de ses collègues de l’université de Lund (Suède) ont permis d’aboutir aux premières découvertes qui allaient collectivement suggérer le rôle de la dopamine comme neurotransmetteur dans le cerveau humain. En août 1957, Montagu a publié le premier article qui démontrait ses découvertes sur les principaux neurotransmetteurs. Dans le cadre de ses recherches, elle a effectué un test sur colonne pour examiner les quantités de noradrénaline, d’adrénaline et de 3-hydroxytyramine dans des tissus extraits du cerveau de plusieurs espèces (rat, lapin, cobaye, poussin, humain et grenouille). Montagu a supposé qu’il pouvait y avoir une autre catécholamine similaire à l’hydroxytyramine, dont elle a confirmé plus tard qu’il s’agissait de la 3,4-dihydroxytyramine (« dopamine ») par chromatographie sur papier des éluats des résines qu’elle a utilisées pour ses tissus cérébraux extraits. En novembre 1957, Carlsson a constaté qu’il pouvait inverser les effets akinétiques que la réserpine induisait chez ses lapins en injectant par voie intraveineuse de la dopamine et du précurseur de la noradrénaline, la 3, 4-dihydroxyphénylalanine (L-DOPA), et a constaté que cela était corrélé à une récupération de la dopamine mais pas de la noradrénaline. Ces données suggéraient que le manque de dopamine pouvait être responsable de l’état akinétique observé chez ses animaux. Le groupe de Carlsson a fini par mettre au point un test permettant de mesurer la concentration de dopamine dans le cerveau et de déterminer où se trouvait la plus forte concentration de dopamine. Ils ont déterminé que la dopamine se trouvait en forte concentration dans le striatum, la plus grande composante des ganglions de la base. À l’époque, il était déjà bien connu que les ganglions de la base jouaient un rôle clé dans les fonctions motrices volontaires. Ces résultats ont contribué à façonner les premières hypothèses selon lesquelles la dopamine pourrait être un neurotransmetteur clé dans le contrôle de la fonction motrice.
Qu’est-ce que la dopamine ? Où est-elle produite dans le cerveau ? Comment est-elle produite ?
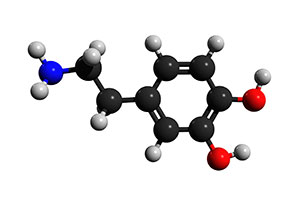
Figure 1 : molécule du neurotransmetteur Dopamine
Depuis sa découverte initiale, la Dopamine, également connue sous le nom de 3, 4-Dihydroxytyramine, a été fortement caractérisée. Elle est constituée d’un noyau benzénique avec deux groupes latéraux hydroxyle attachés à un groupe amine via un groupe éthyle. Elle est produite par les neurones dopaminergiques du cerveau à partir de la tyrosine par l’ajout d’un groupe hydroxyle qui la transforme en L-DOPA (ou Levo-DOPA) et par l’élimination ultérieure d’un groupe acide carboxylique de la chaîne latérale éthyle liée au groupe amine, ce qui donne la Dopamine. Les neurones dopaminergiques qui produisent cette molécule de signalisation sont situés dans le cerveau au niveau de la substantia nigra et de l’aire tegmentale ventrale, toutes deux situées dans le mésencéphale, ainsi que dans le noyau arqué de l’hypothalamus. La dopamine est un neurotransmetteur, c’est-à-dire une substance chimique libérée par les neurones pour transmettre un signal électrique d’un neurone à l’autre afin de transmettre un signal vers et depuis le système nerveux central. Après la production de dopamine, le neurotransmetteur est emballé dans une vésicule synaptique, le transporteur monoamine vésiculaire 2 (VMAT2) et stocké jusqu’à ce que les potentiels d’action induisent la libération de dopamine dans la fente synaptique et provoquent la liaison aux récepteurs de dopamine sur le neurone postsynaptique.
Les neurotransmetteurs de la dopamine se lient à cinq sous-types de récepteurs de la dopamine : D1, D2, D3, D4 et D5, qui sont des membres de la famille des récepteurs couplés aux protéines G qui sont divisés en deux sous-classes principales : D-1-like et D-2-like. La liaison de la dopamine à ces récepteurs déclenche des cascades de signaux responsables de l’activation de fonctions dans les zones du cerveau associées où chaque type de récepteur est le plus répandu. Les récepteurs de type D1 sont plus répandus que les récepteurs de type D2. Pour comprendre comment la dopamine fonctionne dans le cerveau humain en tant que neurotransmetteur, il faut examiner l’effet de la liaison de la dopamine aux types de récepteurs de type D1 et D2 pour exercer leurs effets via des systèmes de second messager. La liaison de la dopamine aux récepteurs de type D1 (D1 et D5) entraîne une excitation via l’ouverture des canaux Na+ ou une inhibition via l’ouverture des canaux K+. La stimulation des récepteurs de type D1 induit l’activation de l’adénylate cyclase, l’enzyme qui convertit l’ATP en AMPc, ce qui augmente les niveaux d’AMPc et entraîne la désinhibition de la protéine kinase A (PKA) qui phosphoryle des cibles en aval telles que la protéine CREB (cAMP regulatory element binding protein). La translocation de la CREB vers le noyau et la transcription de nombreux gènes en fonction de la CREB sont responsables de la plasticité synaptique nécessaire à l’apprentissage et à la formation de la mémoire. En revanche, la liaison des récepteurs de type D-2 (D2, D3 et D4) conduit à l’inhibition du neurone cible en exerçant un effet opposé d’inhibition de l’adénylate cyclase par couplage aux protéines G Gi/o qui diminue la production d’AMPc. Que la dopamine soit excitatrice ou inhibitrice est une question de quel type d’effet sur un neurone cible est exercé qui est basé sur quels types de récepteurs sont sur la surface de la membrane du neurone et comment le neurone répond aux augmentations ou diminutions de la concentration d’AMPc.
Que fait la dopamine dans le cerveau humain ?
La dopamine joue des rôles importants dans la fonction exécutive, le contrôle moteur, la motivation, l’éveil, le renforcement et la récompense par le biais de cascades de signalisation qui s’exercent via la liaison aux récepteurs dopaminergiques au niveau des projections présentes dans la substantia nigra, l’aire tegmentale ventrale et le noyau arqué de l’hypothalamus du cerveau humain.
Dans la substantia nigra, la voie nigro-striatale projette les neurones dopaminergiques de la zone d’entrée (appelée pars compacta) vers le striatum dorsal et joue un rôle primordial dans le contrôle de la fonction motrice et l’apprentissage des habiletés motrices. Si les neurones dopaminergiques de la voie nigro-striatale dégénèrent, cela entraîne un dérèglement du contrôle moteur, caractéristique de la maladie de Parkinson.
Dans l’aire tegmentale ventrale (ATV), la voie mésolimbique se projette du cortex préfrontal vers le noyau accumbens de l’amygdale, le gyrus cingulaire, l’hippocampe et le complexe pyriforme du bulbe olfactif. Les projections dopaminergiques dans l’amygdale et le gyrus cingulaire sont responsables de la formation et du traitement des émotions. Dans l’hippocampe, la présence de neurones dopaminergiques est associée à l’apprentissage, à la mémoire de travail et à la formation de la mémoire à long terme. Enfin, le complexe pyriforme du bulbe olfactif est responsable de l’odorat des humains. Dans la voie mésolimbique, la dopamine est libérée lors de situations agréables, ce qui provoque une excitation et influence le comportement (motivations) à rechercher l’activité ou l’occupation agréable et se lie aux récepteurs dopaminergiques présents dans le noyau accumbens et le cortex préfrontal. L’augmentation de l’activité des projections vers le noyau accumbens joue un rôle majeur dans le renforcement et dans des cas plus extrêmes avec les addictions.
Dans le noyau arqué de l’hypothalamus, les neurones à dopamine constituent la voie tubéro-infundibulaire qui se projette vers l’hypophyse et inhibe la sécrétion de l’hormone prolactine. La dopamine produite par les neurones du noyau arqué est libérée dans les vaisseaux sanguins hypothalamo-hypophysaires qui alimentent l’hypophyse en dopamine pour inhiber la production de prolactine.
Mesure quantitative des niveaux de dopamine dans les échantillons
L’évaluation des niveaux de dopamine en ce qui concerne la dérégulation des fonctions associées à certaines parties du cerveau est une cible attrayante pour la recherche en neurosciences. Enzo Life Sciences propose un kit ELISA de dopamine, qui est un test immunologique compétitif colorimétrique capable de quantifier la dopamine dans le sérum, le plasma, les homogénats de tissus et d’autres fluides biologiques. Ce kit est hautement spécifique de la dopamine humaine et présente une réactivité croisée négligeable entre la détection de la dopamine humaine et de ses analogues. Cet essai immunologique très sensible a une gamme de détection qui peut détecter aussi peu que 1,56 ng/ml et autant que 100ng/ml (Figure 1). En outre, ce produit a été conçu avec une sensibilité élevée, une reproductibilité lot à lot élevée et un faible temps de résultat et offre un protocole simple qui peut produire des résultats fiables et quantitatifs pour nos utilisateurs finaux en moins de 2 heures pour un maximum de 40 échantillons en double.
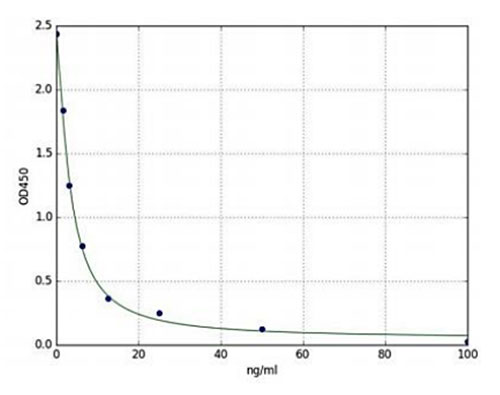
Figure 2 : Courbe standard du kit ELISA Dopamine (ENZ-KIT188) représentant une courbe standard typique (1,56 ng/ml-100ng/ml).
Enzo Life Sciences fournit une grande variété de produits pour vos besoins de recherche en neurosciences et en immunologie. Nous proposons d’autres immunodosages de neurotransmetteurs tels que notre kit ELISA de sérotonine et notre kit ELISA d’histamine, ainsi qu’une large sélection d’anticorps pour étudier les neurotransmetteurs tels que l’ACTH, l’ANP, le BNP, la CCK, le CGRP, le NPY, le GABA, le GLP-1 et la substance P. Le portefeuille complet d’Enzo comprend notre bibliothèque de neurotransmetteurs SCREEN-WELL, qui contient 661 ligands de récepteurs CND dans un format de 96 puits. Veuillez consulter nos plateformes Neuroscience et Signalisation cellulaire/Transduction du signal pour plus d’informations ou n’hésitez pas à contacter notre service d’assistance technique pour toute assistance supplémentaire.