ADVERTISSEMENTS:
Dans cet article, nous allons discuter de Paramecium Caudatum:- 1. Habitude, habitat et culture de Paramecium Caudatum 2. Structure de Paramecium Caudatum 3. Locomotion 4. Nutrition 5. Respiration et excrétion 6. Osmorégulation 7. Comportement 8. La reproduction 9. Comportement aberrant dans la reproduction 10. Quelques particules cytoplasmiques.
Contenu :
- Habitat, Habitat et culture de la paramécie caudatum
- Structure de la paramécie caudatum
- Locomotion de la paramécie caudatum
- Nutrition de la paramécie caudatum
- Respiration et excrétion de la paramécie caudatum
- Osmorégulation de la paramécie caudatum
- Comportement du Paramecium Caudatum
- Reproduction dans le Paramecium Caudatum
- Comportement aberrant dans la reproduction dans le Paramecium Caudatum
- Certaines particules cytoplasmiques signalées dans le Paramecium Caudatum
. Paramecium Caudatum
- Habitat, Habitat et culture de la Paramecium Caudatum :
- Culture de la paramécie :
- Structure de la Paramecium Caudatum :
- (i) Taille et forme de la Paramecium Caudatum :
- (ii) Pellicule :
- (iii) Cils :
- (iv) Système infra-ciliaire :
- (v) Sillon buccal et cytopyge :
- (vi) Cytoplasme :
- (vii) Ectoplasme :
- (viii) Trichocystes :
- (ix) Neuromotorium et fibrilles associées :
- (x) Endoplasme :
- (xi) Vacuoles contractiles :
- (xii) Vacuoles alimentaires :
- Locomotion de la paramécie caudatum :
- (i) Métabolie ou contorsions corporelles :
- (ii) Locomotion ciliaire :
- Nutrition de la Paramécie Caudatum :
- Mécanisme d’alimentation :
- Digestion et Egestion du Paramecium Caudatum :
- Respiration et excrétion de Paramecium Caudatum :
- Comportement de Paramecium Caudatum :
- (i) Réactions au contact (Thigmotaxie) :
- (ii) Réactions aux produits chimiques (chimiotaxie) :
- (iii) Réactions à la température (Thermotaxie) :
- (iv) Réactions à la lumière (Phototaxie) :
- (v) Réactions au courant électrique (Galvanotaxie) :
- (vi) Réactions au courant d’eau (Rhéotaxie) :
- (vii) Réactions à la gravité (Géotaxie) :
- Reproduction chez la Paramecium Caudatam :
- (i) Fission binaire transverse :
- (ii) Conjugaison:
- Comportement aberrant dans la reproduction chez Paramecium Caudatum :
- (i) Endomixie :
- (ii) Autogamie :
- (iii) L’hémixie :
- (iv) La cytogamie :
- Certaines particules cytoplasmiques signalées dans le Paramecium Caudatum :
- (i) Particules kappa :
- (ii) mµ, Particules :
- (iii) Particules Pi :
- (iv) Particules Lambda :
Habitat, Habitat et culture de la Paramecium Caudatum :
Paramecium caudatum (Gr., paramekes = oblong ; L., caudata = queue) se trouve couramment dans les étangs d’eau douce, les mares, les fossés, les ruisseaux, les lacs, les réservoirs et les rivières. On la trouve surtout en abondance dans les étangs stagnants riches en matières en décomposition, dans les infusions organiques et dans les eaux d’égout. Paramecium caudatum est un organisme vivant librement et cette espèce a une distribution mondiale.
Culture de la paramécie :
ADVERTISSEMENTS:
Prenez des herbes submergées dans un étang et placez-les dans un bocal d’eau distillée, couvrez le bocal et laissez-le pourrir ; des essaims de Paramecia apparaîtront en quelques jours. Faites maintenant bouillir du foin dans de l’eau, décantez l’infusion et ajoutez quelques grains de blé, et laissez reposer jusqu’à ce qu’elle soit trouble de bactéries.
Transférez les Paramecia du premier bocal dans ce liquide où ils se multiplieront rapidement. Les infusions de foin seules produiront des Paramecia montrant la présence de kystes, et des kystes ressemblant à des grains de sable ont été signalés, mais il n’y a pas de preuve que les Paramecia forment des kystes, car ils n’ont jamais été confirmés.
Structure de la Paramecium Caudatum :
(i) Taille et forme de la Paramecium Caudatum :
La Paramecium caudatum (Fig. 20.1) est un organisme microscopique et visible à l’œil nu comme un minuscule corps allongé.
ADVERTISSEMENTS:
Il apparaît gris clair ou blanc mesurant couramment entre 170 et 290 microns de longueur et peut atteindre une longueur allant jusqu’à 300-350 microns. P. caudatum ressemble à la semelle d’une pantoufle ou d’une chaussure, d’où le nom d’animalcule de la pantoufle. Il est quatre fois plus long que large et quelque peu cylindrique avec des extrémités nettement différentes.
La partie antérieure, qui se déplace vers l’avant, est mince avec une extrémité émoussée ou arrondie, tandis que l’extrémité postérieure est quelque peu pointue ou conique. La partie la plus large de l’organisme se situe juste en dessous du milieu. Le corps de l’animal est de forme asymétrique montrant une surface orale ou ventrale bien définie et une surface aborale ou dorsale.
(ii) Pellicule :
Le corps est recouvert d’une pellicule fine, à double couche, élastique et ferme, faite de gélatine. La pellicule maintient la forme de l’animal mais est suffisamment élastique pour permettre des contractions. La pellicule possède une double membrane, la membrane externe est en continuité avec les cils et la membrane interne avec l’ectoplasme. Sous le plus fort grossissement du microscope, la pellicule montre des dépressions rectangulaires ou hexagonales sur sa surface.

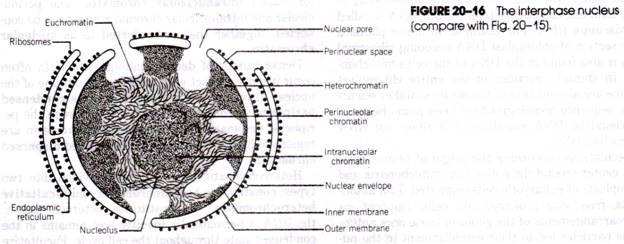
Cette disposition est préservée sur la surface dorsale de la Paramecium mais sur la surface ventrale les crêtes convergent devant et derrière vers une ouverture préorale et postorale. Chaque dépression hexagonale est perforée par une ouverture centrale à travers laquelle émerge un seul cilium. Les marges antérieures et postérieures des dépressions hexagonales portent les ouvertures des trichocystes.
L’étude au microscope électronique de la pellicule (Fig. 20.2) par Ehret et Powers (1957) a révélé que les dépressions hexagonales correspondent à des séries régulières de cavités, les alvéoles. Toutes les alvéoles forment collectivement une couche alvéolaire continue, qui est délimitée par une membrane alvéolaire externe et une membrane alvéolaire interne.
La couche externe se trouve en contact étroit sous la membrane cellulaire externe. Par conséquent, la pellicule comprend la membrane cellulaire externe, la membrane alvéolaire externe et la membrane alvéolaire interne.
(iii) Cils :
L’ensemble du corps est recouvert de nombreuses petites projections semblables à des cheveux, appelées cils. Les cils se produisent en rangées longitudinales sur tout le corps, cette condition est connue comme holotriche dans laquelle les cils du corps sont égaux. Les cils ont la même structure que les flagelles, ils ont une gaine protoplasmique externe ou une membrane plasmique avec neuf doubles fibrilles longitudinales dans un anneau périphérique. Dans certains cils, les neuf fibrilles extérieures ne sont pas appariées.
Avertissements:
Il y a deux fibrilles longitudinales centrales qui sont plus fines que les fibrilles externes. Chaque cilium naît d’un granule basal ou kinétosome. Les neuf paires de fibrilles périphériques fusionnent pour former la paroi du kinétosome, ainsi, le kinétosome est un tube ouvert ou fermé à son extrémité inférieure, les deux fibrilles centrales s’arrêtent au niveau de la pellicule chez la plupart des ciliés.
Sortant du kinétosome se trouve un mince rhizoplaste qui ne rejoint pas le noyau. De nombreux Métazoaires possèdent également des cils, leur structure est la même, sauf que le granule basal est différent et qu’il possède de fins filaments ou des fibres d’enracinement s’étendant vers le bas dans le cytoplasme. Mais les cils diffèrent des flagelles en étant généralement plus nombreux et de taille plus courte.
La ciliature peut être divisée de manière pratique en cils corporels ou somatiques qui se trouvent à la surface du corps, et en cils oraux qui sont associés à la région buccale. Les cils du corps sont égaux mais ils sont plus longs à l’extrémité postérieure, d’où le nom de caudatum. Les cils sont des organites de locomotion et de collecte de nourriture, ils agissent également comme des récepteurs sensoriels et détectent les stimuli du milieu extérieur.

Ultra Structure des cils :
Les cils et les flagelles ont une composition fibrillaire. A la base, le cilium a un diamètre d’environ 0,2 micron ou 2 000 A0 qui peut se trouver jusqu’à 10 microns au-dessus de la surface de la cellule. Les cils sont délimités par une membrane unitaire de 90 A0 d’épaisseur qui ressemble à la membrane plasmique et reste en continuité avec elle. L’espace délimité du cilium contient une substance aqueuse appelée matrice.
Dans la matrice, restent enchâssées onze fibrilles longitudinales ou microtubules. Sur les onze fibrilles, deux sont situées au centre, tandis que les neuf fibrilles restantes restent disposées de manière périphérique autour des fibrilles centrales. Chacune des neuf fibrilles externes a un diamètre de 360A0 et est composée de deux sous-fibrilles de 180 à 250A0 de diamètre.
Ces sous-fibrilles sont désignées comme la sous-fibrille A et la sous-fibrille B.
Avertissements:
La sous-fibrille A est légèrement plus grande que la sous-fibrille B. La sous-fibrille A donne deux projections épaisses ou bras de son côté. Les bras de la sous-fibrille A de toutes les fibrilles extérieures restent dirigés dans le sens des aiguilles d’une montre. En outre, la sous-fibrille A se trouve plus près du centre du cilium que la sous-fibrille B. Les deux sous-fibrilles ont une paroi commune de 50A0 d’épaisseur.
Les deux fibrilles centrales n’ont pas de sous-fibrilles appariées comme les neuf fibrilles périphériques mais chacune ne contient qu’un seul tubule. Chaque fibrille centrale a un diamètre d’environ 250A0 et est composée d’une paroi de 60A0 d’épaisseur.
Les deux fibrilles centrales restent séparées par un espace de 350A0 et restent enveloppées dans une gaine commune. Gibbnos (1967) a signalé que la gaine des fibrilles centrales donne neuf liens ou rayons orientés radialement à chaque sous-fibrille A.

La microscopie électronique à haute résolution a révélé que chacune des fibrilles périphériques et centrales des cils et des flagelles est composée de dix à douze filaments de 40A0 d’épaisseur. Chaque filament est perlé. Chaque perle reste disposée en réseau de 40 par 50A0 dans le plan de la paroi du tubule. Ces perles sont considérées comme la sous-unité de base de la structure du tubule.
(iv) Système infra-ciliaire :
Le système infra-ciliaire est situé juste en dessous des alvéoles pelliculaires. Il est constitué du kinétosome ou corps basal et du kinétodesme. Les cils naissent des kinétosomes et de chaque kinétosome naît une délicate fibrille cytoplasmique appelée kinétodesme, (Fig. 20.2).
Situé sous la pellicule légèrement à droite, mais joint à tous les cinétosomes d’une rangée longitudinale, se trouve un faisceau longitudinal de plusieurs cinétodesmates, un cinétodesme de chaque cinétosome s’étend sur une distance antérieurement dans son propre faisceau de cinétodesmates.
Une rangée longitudinale de cinétosomes avec leurs cinétodesmates forme une unité longitudinale appelée cinétie. Tous les kinety ou kinetia constituent le système infra-ciliaire d’un cilié. Les kinety se trouvent dans le cortex sous la pellicule, leur nombre est assez constant pour chaque cilié.
Le système infra-ciliaire contrôle et coordonne les mouvements des cils, et il entraîne la formation d’organites lors de la division cellulaire, par exemple, certains kinetia forment la bouche. Dans la fission binaire des ciliés, les cinéties sont coupées transversalement en deux, chacune allant à une cellule fille, c’est la fission péricinétale.
(v) Sillon buccal et cytopyge :
Sur le côté ventrolatéral se trouve une grande dépression oblique et peu profonde appelée sillon oral ou péristome qui donne à l’animal un aspect asymétrique. Il s’étend obliquement vers l’arrière d’un côté (généralement de gauche à droite mais dans certains cas de droite à gauche) et se termine un peu derrière le milieu du corps. Le sillon oral débouche sur une courte dépression conique en forme d’entonnoir appelée vestibule.
ADVERTISSEMENTS:
Le vestibule débouche directement sur l’ouverture fixe de forme ovale appelée cytostome (bouche). S’étendant directement du cytostome vers le centre du corps se trouve le large cytopharynx. Le cytopharynx tourne ensuite brusquement vers le côté postérieur pour devenir l’œsophage effilé et mince.
Donc, l’œsophage est à peu près parallèle à la surface du corps de la Paramécie, sauf à son extrémité postérieure. Là, l’œsophage se tourne à nouveau vers le centre de l’animal pour déboucher dans la vacuole alimentaire en formation.
Le cytopyge (également appelé anus cellulaire ou tache anale ou cytoprocte) se trouve sur la surface ventrale du corps presque verticalement derrière le cytostome ou la bouche. Les particules alimentaires non digérées sont éliminées par le cytopyge. La ciliation du cytopharynx est très compliquée.
Gelei (1934) a signalé la présence de quatre rangées et Lund (1941) a observé au moins quatre rangées. Une structure appelée penniculus se trouve sur la paroi gauche du cytopharynx et s’enroule en spirale sur environ 90 degrés de sorte que son extrémité postérieure se trouve sur la surface orale (ventrale) de l’œsophage.
Selon Lund, le penniculus est composé de huit rangées de cils disposés en deux blocs rapprochés de chacun. Une bande similaire composée de quatre rangées de longs cils, moins compacts que dans le penniculus, est appelée quadrulus. Elle descend en spirale le long de la paroi dorsale de la cavité buccale et se termine près du penniculus.
Le penniculus et le quadrulus ont été appelés à tort une membrane ondulée par certains travailleurs. Le quadrulus et le penniculus contrôlent le passage de la nourriture. On ne sait pas comment les cils fonctionnent, probablement leurs fibrilles se contractent de manière rythmique ce qui provoque la flexion. Gelei (1925) a souligné que la fonction du penniculus est le forçage des éléments alimentaires dans le corps.

(vi) Cytoplasme :
Le cytoplasme se différencie en une zone étroite, externe ou corticale, appelée ectoplasme et une région plus large, interne ou médullaire, appelée endoplasme.
(vii) Ectoplasme :
L’ectoplasme (ectosarc ou cortex) est une partie permanente du corps, délimitée de façon frappante de l’endoplasme. L’ectoplasme forme une couche externe ferme, claire, fine et dense. Il contient les trichocystes, les cils et les structures fibrillaires et est délimité extérieurement par une couverture appelée pellicule.
(viii) Trichocystes :
Enchâssés dans l’ectoplasme à angle droit de la surface se trouvent de petits sacs fusiformes appelés trichocystes. Un petit point sur chaque marge antérieure et postérieure de l’hexagone marque la position d’un trichocyste. Ils sont remplis d’un fluide réfringent et dense ayant une substance gonflante, à l’extrémité extérieure se trouve une tête ou un pic conique.
Les trichocystes se trouvent perpendiculaires dans l’ectoplasme, ils s’ouvrent par de petits pores sur les crêtes des zones hexagonales de la pellicule.
Ils naissent des kinétosomes des cils, puis migrent et se localisent à égale distance dans l’endoplasme. Lorsque l’animal est irrité, les trichocystes sont déchargés sous forme de longs fils collants. Un trichocyste déchargé possède une pointe opaque semblable à un clou inversé, et une longue tige striée, mais la tige n’est pas visible à l’état non déchargé et se forme probablement pendant la décharge.
La fonction des trichocystes est incertaine, mais ils sont déchargés en réaction aux contacts locaux et aux blessures, ils peuvent servir d’organites de défense.
Mais cette fonction est incertaine car les trichocystes sont inefficaces contre Didinium, le principal prédateur de la Paramécie, ils peuvent servir à fixer l’animal à un endroit pendant l’alimentation. Chez certains ciliés, les trichocystes agissent comme des organelles d’infraction. Après la décharge des trichocystes, régénérés à partir des kinétosomes.


(ix) Neuromotorium et fibrilles associées :
Selon Lund (1933) sur la paroi dorsale gauche du cytopharynx, à peu près au niveau de la marge postérieure du cytostome, se trouve une très petite masse bilobée, le neuromotorium. A partir du neuromotorium, des fibrilles rayonnent dans l’endoplasme.
D’entre elles, quatre ou plus passent généralement presque jusqu’à la paroi dorsale du corps, mais les autres sont plus courtes et leur position n’est pas définie. Toutes sont appelées fibrilles endoplasmiques. Leur fonction est inconnue, mais elles peuvent coordonner les mouvements d’alimentation des cils oraux. Les fibrilles peuvent également donner un support mécanique, une élasticité, une contractilité, une conductivité et une influence métabolique.

(x) Endoplasme :
L’endoplasme ou médulla est la partie la plus fluide et la plus volumineuse du cytoplasme qui contient de nombreux granules cytoplasmiques ainsi que d’autres inclusions et structures de nature spécialisée. Les inclusions cytoplasmiques sont des mitochondries, des appareils de Golgi, des vacuoles, des cristaux, des granules et des chromidies, etc. D’autres structures, à savoir les noyaux, les vacuoles contractiles et les vacuoles alimentaires se trouvent également dans l’endoplasme.
Nucléus :
Dans l’endoplasme près du cytostome se trouvent deux noyaux, c’est-à-dire que la paramécie est hétérocaryote, un grand macronoyau ellipsoïde et granuleux et d’autres petits micronoyaux compacts.
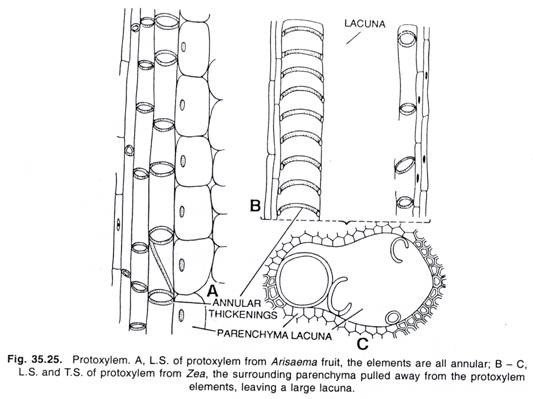
Le macronoyau est un corps ostensible, ellipsoïdal ou réniforme. Il est de type compact contenant des fils fins et des granules chromatiniens discrets serrés de taille variable et noyés dans une matrice achromatique. Il possède de nombreux nucléoles et beaucoup plus de matériel chromatinien (ADN).
C’est un noyau somatique ou végétatif. Il se divise de façon amitotique et contrôle les fonctions végétatives (activités métaboliques) de l’animal. Il ne subit pas de mitose.
Le micronoyau est petit, compact et sphérique. Il se trouve généralement à proximité du macronoyau souvent dans une concavité. De fins granules et fils de chromatine sont uniformément répartis dans la structure. Le micronoyau se divise mitotiquement et contrôle la reproduction.
Le nombre de micronoyaux varie selon les espèces ; il est de un chez P. caudatum, de deux chez P. aurelia et de plusieurs chez P. multimicronucleatum. Le micronoyau contient un nucléole distinct chez P. aurelia mais on ne le trouve pas chez P. caudatum. Moses (1949 ; 1950) a signalé que le macronoyau et le micronoyau sont identiques dans leur composition chimique.
(xi) Vacuoles contractiles :
Il y a deux grandes vacuoles contractiles remplies de liquide, chacune située près d’une extrémité du corps près de la surface dorsale. Leur position est fixe (contrairement aux Amibes), elles se situent entre l’ectoplasme et l’endoplasme, mais ce sont des organites temporaires qui disparaissent périodiquement. Chez certaines espèces, ils semblent avoir une membrane de revêtement, auquel cas ils ne disparaissent pas entièrement pendant la systole.
Connectés à chaque vacuole contractile se trouvent cinq à douze canaux radiaires tubulaires, chacun étant constitué d’une partie terminale, d’une longue ampoule qui s’affaisse lorsqu’elle est vide, et d’un court canal injecteur qui s’ouvre dans la vacuole.

Les canaux communiquent avec une grande partie du corps d’où ils captent les liquides et les versent dans la vacuole qui est, ainsi, reconstituée et augmente de taille, lorsque la vacuole contractile atteint sa taille maximale, elle se contracte brusquement (systole) et décharge son contenu par un pore permanent de la pellicule, puis les canaux forment à nouveau les vacuoles contractiles, les canaux ne disparaissent pas entièrement car ce sont des structures permanentes.
Les deux vacuoles contractiles se déchargent irrégulièrement, la postérieure se contracte plus rapidement car elle est proche du cytopharynx et plus d’eau y entre. La fonction principale des canaux et des vacuoles contractiles est hydrostatique, ils éliminent l’excès d’eau du protoplasme, l’eau est en partie absorbée et en partie prise en charge lors de l’alimentation.
Les déchets azotés comportent des composés ammoniacaux et quelques urates qui sont expulsés des vacuoles contractiles en même temps que le CO2 mais il n’y a pas de preuve que la matière excrétoire soit sécrétée par le protoplasme dans les canaux ou les vacuoles contractiles.
Il y a un cytopyge permanent fixe ou anus sous le péristome à un côté du cytopharynx, par lequel les restes non digérés de nourriture et les urates sont expulsés.
L’étude au microscope électronique des vacuoles contractiles a révélé que chaque appareil contractile est constitué de certains des tubules du réticulum endoplasmique, des tubules néphridiens, des canaux nourriciers, des vacuoles accessoires (canaux radiaux) et de la vacuole contractile principale. Les vacuoles accessoires sont supposées être les ampoules des canaux nourriciers (figure 20.11).
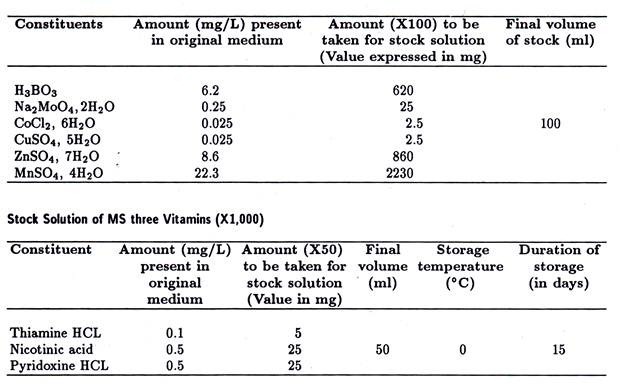
(xii) Vacuoles alimentaires :
Ce sont des corps grossièrement sphériques, non contractiles, variant en taille et en nombre, situés dans l’endoplasme. Ils contiennent des particules alimentaires ingérées, principalement des bactéries, et une petite quantité de liquide délimité par une fine membrane définie. Volkonsky (1934) a proposé le nom de gastrioles pour ces vacuoles. Les granules digestifs sont associés aux vacuoles alimentaires.
Locomotion de la paramécie caudatum :
La paramécie caudatum effectue une locomotion par deux méthodes, à savoir la métabolie ou les contorsions du corps et par les cils.
(i) Métabolie ou contorsions corporelles :
Le corps de la Paramecium Caudatum possède une élasticité, elle peut se serrer dans un passage plus étroit que son corps, après quoi le corps reprend sa forme normale. Ce changement temporaire de la forme du corps est le métabole, il est provoqué chez la Paramecium par le protoplasme.
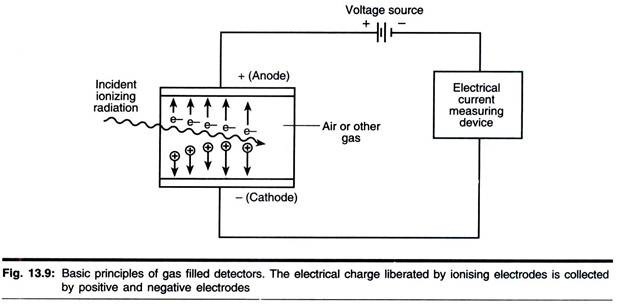
(ii) Locomotion ciliaire :
La locomotion apportée par les cils est la principale méthode. Les cils peuvent battre en avant ou en arrière permettant à l’animal de nager vers l’avant ou vers l’arrière.
Normalement, l’animal nage vers l’avant, les cils battant vers l’arrière mais obliquement, les cils se raidissent et se courbent rapidement vers l’arrière pour presque toucher la surface du corps, c’est ce qu’on appelle la course efficace ; puis les cils deviennent mous et reviennent lentement à la position verticale initiale, c’est , la course de récupération.
Les cils d’une même rangée transversale battent ensemble et ceux d’une même rangée longitudinale battent l’un après l’autre de l’extrémité antérieure à l’extrémité postérieure.

Ce mouvement coordonné des cils est appelé rythme métachronique, qui est dû au système infra-ciliaire ; cela provoque la nage en avant de l’animal. Mais lorsque les cils du corps battent obliquement vers l’arrière, alors, en même temps, les cils plus longs du sillon oral battent plus vigoureusement, ce qui provoque une déviation de l’extrémité antérieure vers la gauche.
L’action des cils du corps et du sillon oral fait tourner l’animal sur son grand axe. Cette rotation se fait toujours vers la gauche (sauf chez P. calkinsi qui tourne en spirale vers la droite).
Cette combinaison de mouvement vers l’avant, d’embardées et de rotation fait que l’animal se déplace vers l’avant dans une trajectoire en spirale dans le sens inverse des aiguilles d’une montre. Ce chemin a un axe droit, et la même surface corporelle de l’animal reste vers l’axe du chemin en spirale. Mais en nageant à reculons, toutes les espèces tournent vers la droite.
Le battement ciliaire peut être inversé de sorte que les cils se déplacent obliquement vers l’avant par lequel l’animal nage à reculons. Par l’action ciliaire, la paramécie se déplace avec une vitesse de 1500 microns ou même plus par seconde.
Jennings a soutenu que la spirale de la Paramécie est due au fait que, tandis que les cils frappent principalement vers l’arrière, ils le font obliquement vers la droite, ce qui fait que l’animal roule vers la gauche.
De plus, cette déviation du corps vers la surface aborale est due en grande partie à la plus grande puissance de la course effective des cils oraux qui frappent plus directement vers l’arrière. Le résultat – la rotation de la paramécie sur son axe long – permet ainsi à la paramécie de suivre une trajectoire plus ou moins rectiligne en formant de grandes spirales.
Nutrition de la Paramécie Caudatum :
Dans la Paramécie Caudatum, la nutrition est holozoïque. La nourriture comprend principalement des bactéries et des protozoaires minuscules. La Paramecium n’attend pas la nourriture mais la chasse activement.
On prétend que la Paramecium Caudatum montre un choix dans la sélection de sa nourriture, mais il ne semble pas y avoir de base pour cela bien qu’elle engloutisse seulement certains types de bactéries ; les données disponibles suggèrent que 2 à 5 millions d’individus de Bacillus coli sont dévorés par une seule Paramecium en 24 heures. Elle se nourrit également de plantes unicellulaires comme les algues, les diatomées, etc. et de petits morceaux d’animaux et de légumes.
Mécanisme d’alimentation :
Lorsque la Paramecium Caudatum entre dans une région où la nourriture est abondante, elle vient se reposer. Elle ne se nourrit qu’au repos ou lorsqu’elle nage très lentement, elle ne se nourrit jamais lorsqu’elle nage rapidement. Le battement des cils du sillon buccal provoque un tourbillon conique d’eau chargée de nourriture qui est balayé dans le sillon buccal à une distance en avant de l’extrémité antérieure (figure 20.16).
Les particules de nourriture se dirigent ensuite vers le vestibule d’où certaines particules alimentaires sont rejetées et expulsées, mais d’autres passent dans le cytostome.
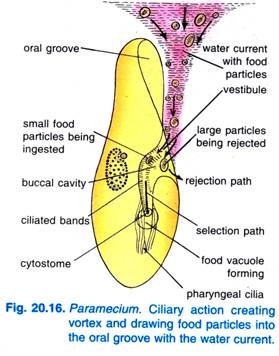
À l’extrémité du cytopharynx, se forme une vacuole alimentaire qui se remplit de particules d’aliments. Le quadrulus et le peniculi contrôlent le passage des aliments dans la vacuole alimentaire qui se forme latéralement. Lorsque la vacuole alimentaire atteint une certaine taille, les fibres post-buccales enserrent la vacuole alimentaire qui est pincée par elles et reprend sa course.
La vacuole contient un peu d’eau en plus de la nourriture. Des mouvements rotatifs de streaming de l’endoplasme appelés cyclosis transportent les vacuoles alimentaires le long d’un parcours défini qui est fonctionnellement équivalent à un tube digestif.
Le tractus part de l’extrémité du cytopharynx, puis vers la face postérieure, puis vers l’avant pour circuler avec l’endoplasme, puis vers la face dorsale, puis vers l’extrémité antérieure, puis vers le bas jusqu’au cytopyge. Au début de son voyage, la vacuole alimentaire diminue de taille, puis augmente à nouveau.

Digestion et Egestion du Paramecium Caudatum :
Pendant la cyclosis, la digestion se fait par des enzymes sécrétées par le protoplasme dans les vacuoles. Lors de la digestion, les protéines sont transformées en acides aminés, les glucides en sucres solubles et en glycogène, et les graisses sont probablement aussi digérées.
Le contenu des vacuoles alimentaires est d’abord acide (pH d’environ 4) puis devient alcalin, la digestion majeure a lieu pendant la phase alcaline. Les matières non digérées sont éjectées à travers le cytopyge avec une certaine force.
La cyclosis peut être démontrée expérimentalement ; si du lait coloré au rouge Congo est donné à manger à la paramécie, les globules gras du lait dans les vacuoles alimentaires deviendront d’abord rouges à cause de la réaction acide des enzymes, puis ils passeront des nuances de violet au bleu à cause de la réaction alcaline, les vacuoles montreront le déroulement de la cyclosis.
Respiration et excrétion de Paramecium Caudatum :
L’échange de gaz (oxygène et dioxyde de carbone) se fait à travers la pellicule semi-perméable comme les autres protozoaires d’eau douce par le processus de diffusion. La Paramecium Caudatum obtient son oxygène de l’eau environnante. Le dioxyde de carbone et les déchets organiques comme l’ammoniac résultant du métabolisme sont probablement excrétés en diffusant vers l’extérieur dans l’eau en sens inverse.

6. Osmorégulation chez Paramecium Caudatum :
Paramecium Caudatum possède deux vacuoles contractiles, une antérieure et une postérieure. La fonction des vacuoles contractiles est l’osmorégulation, c’est-à-dire la régulation du contenu en eau du corps et peut servir également à l’excrétion des déchets azotés comme l’urée et l’ammoniac.
L’excès d’eau (à cause de l’endosmose continue) au sein du cytoplasme est sécrété dans les tubules du réticulum endoplasmique et se dirige vers les tubules néphridiens → canaux nourriciers → et se rassemblent dans l’ampoule d’une série de 6 à 11 canaux rayonnants qui convergent vers et se déchargent dans chaque vacuole. Les canaux sont plus apparents lorsqu’une vacuole se forme.
Lorsque chaque vacuole est gonflée (diastole) à une certaine taille, elle se contracte (systole) et se décharge à l’extérieur probablement par un pore. Les vacuoles contractiles se contractent alternativement, à des intervalles de 10 à 20 secondes.
La vacuole contractile postérieure fonctionne plus rapidement que la vacuole antérieure en raison de l’admission d’une grande quantité d’eau dans la région postérieure par le cytopharynx. Les vacuoles contractiles maintiennent une concentration optimale d’eau dans le cytoplasme du corps en éliminant l’excès.

Comportement de Paramecium Caudatum :
Les réponses de Paramecium Caudatum à divers types de stimuli sont apprises par l’étude de ses réactions et du regroupement ou de la dispersion des individus dans une culture. La réponse est positive si l’animal se rapproche d’un stimulus et négative lorsqu’il s’en éloigne. A un stimulus défavorable, l’animal continue à donner la réaction d’évitement jusqu’à ce qu’il s’échappe.
Dans la réaction d’évitement, le battement ciliaire s’inverse, l’animal recule sur une courte distance, puis tourne sur une trajectoire conique en faisant pivoter l’extrémité antérieure aboralement tout en pivotant sur la pointe postérieure. Tous les ajustements sont effectués par essais et erreurs. Des expériences ont montré que l’extrémité antérieure de l’animal est plus sensible que les autres parties.
Les réponses de la paramécie à différents stimuli peuvent être regroupées comme suit :
(i) Réactions au contact (Thigmotaxie) :
La réponse au contact est variée chez la Paramécie. Si l’extrémité antérieure est légèrement touchée avec une pointe fine, une forte réaction d’évitement se produit. Lorsqu’une Paramecium qui nage entre en collision avec un objet dans l’eau, mais si on la touche ailleurs, il peut n’y avoir aucune réaction. Un individu qui se déplace lentement répond souvent, positivement au contact avec un objet en venant se poser sur lui.
(ii) Réactions aux produits chimiques (chimiotaxie) :
Généralement, les Paramecia répondent à un stimulus chimique en évitant la réaction. Si une goutte de solution saline faible (0,5 pour cent) est introduite dans une population de Paramecium sur une microglissière, les animaux répondent par la réaction d’évitement et aucun n’entre dans la goutte. En revanche, pour les acides, la réponse est positive même lorsque la concentration est suffisamment forte pour les tuer.
(iii) Réactions à la température (Thermotaxie) :
La paramécie recherche une température optimale de 24 à 28°C. Lorsqu’un changement de température se produit de façon marquée au-dessus ou au-dessous de la plage optimale, les Paramecia montrent une réaction d’évitement. Une chaleur plus importante stimule des mouvements rapides et des réactions d’évitement jusqu’à ce que les animaux s’échappent ou soient tués.
(iv) Réactions à la lumière (Phototaxie) :
À l’exception de la Paramecium bursaria verte, qui est positivement phototactique, les autres espèces sont indifférentes à la lumière ordinaire. Cependant, lorsque l’intensité lumineuse est soudainement et fortement augmentée, une réaction négative s’ensuit généralement. Les paramécies présentent une réaction négative immédiate aux rayons ultraviolets.
(v) Réactions au courant électrique (Galvanotaxie) :
Les paramécies répondent à des stimuli électriques. Lorsque deux électrodes sont placées en face l’une de l’autre dans un plat peu profond contenant des Paramecia et qu’un courant constant est appliqué, tous les organismes nagent dans la même direction vers la cathode ou électrode négative où ils se concentrent en grand nombre.
Si le sens du courant électrique est inversé pendant que les Paramecia nagent vers la cathode, les organismes inversent le sens et nagent vers la nouvelle cathode.

(vi) Réactions au courant d’eau (Rhéotaxie) :
Les paramécies présentent une rhéotaxie positive. Dans un courant d’eau doux, les Paramecia se déplacent le plus souvent avec le courant, avec leurs extrémités antérieures en amont.
(vii) Réactions à la gravité (Géotaxie) :
Les Paramecia présentent généralement une réponse négative à la gravité comme on peut le voir dans une culture où de nombreux individus se rassemblent près sous le film de surface avec leurs extrémités antérieures vers le haut. Si les paramécies sont introduites dans un tube en forme de U inversé rempli d’eau et bouché aux deux extrémités, elles se déplacent immédiatement vers le haut dans la partie horizontale du tube.
Reproduction chez la Paramecium Caudatam :
Paramecium Caudatum se reproduit asexuellement par fission binaire transverse et subit également plusieurs types de réorganisation nucléaire, comme la conjugaison, l’endomixie, l’autogamie, la cytogamie et l’hémixie, etc.
(i) Fission binaire transverse :
La fission binaire transverse est le type de reproduction asexuée le plus courant chez la Paramécie. C’est un processus asexué tout à fait unique dans lequel un spécimen pleinement développé se divise en deux individus filles sans laisser de cadavre parental.
Le plan de division passe par le centre de la cellule et dans un plan à angle droit par rapport au grand axe du corps. La division du corps cellulaire dans son ensemble est toujours précédée par la division des noyaux ; il semble en effet que la reproduction soit initiée par l’activité et la division nucléaires.

Paramecium Caudatum se reproduit par fission binaire transversale pendant les conditions favorables. Dans la fission binaire, le micronoyau se divise par mitose en deux micronoyaux filles, qui se déplacent vers les extrémités opposées de la cellule. Le macronoyau s’allonge et se divise transversalement par amitose.
Un autre cytopharynx est bourgeonné et deux nouvelles vacuoles contractiles apparaissent, une près de l’extrémité antérieure et une autre près de l’extrémité postérieure. Pendant ce temps, un sillon de constriction apparaît près du milieu du corps et s’approfondit jusqu’à ce que le cytoplasme soit complètement divisé.
Les deux paramécies « filles » résultantes sont de taille égale, chacune contenant un ensemble d’organelles cellulaires. Des deux paramécies filles produites, celle qui est antérieure est appelée proter et celle qui est postérieure est appelée opisthe. Elles atteignent leur taille maximale avant qu’une autre division ne se produise.
Le processus de fission binaire nécessite environ deux heures pour se terminer et peut se produire de une à quatre fois par jour, donnant lieu à 2 à 16 individus. Environ 600 générations sont produites en un an.
Le taux de multiplication dépend des conditions externes de nourriture, de température, d’âge de la culture et de densité de population ; également des facteurs internes d’hérédité et de physiologie. Naturellement, si tous les descendants d’un individu survivaient et se reproduisaient, le nombre de paramécies produites serait bientôt égal au volume de la terre.
Le terme clone est utilisé pour désigner l’ensemble des individus qui ont été produits à partir d’un individu par fission. Tous les membres d’un clone sont héréditairement semblables.
(ii) Conjugaison:
Ordinairement, la Paramecium Caudatum se multiplie par fission binaire pendant de longues périodes, mais par intervalles, cela peut être interrompu par la réunion de deux animaux le long de leurs surfaces orales pour le processus sexuel de conjugaison.
La conjugaison est définie comme l’union temporaire de deux individus qui échangent mutuellement du matériel micro nucléaire. C’est un type unique de processus sexuel dans lequel deux organismes se séparent peu après l’échange de matériel nucléaire.
Sonneborn (1947), sur la base du comportement d’accouplement de Paramecium Caudatum, a rapporté que chaque espèce de Paramecium existe dans un certain nombre de variétés ou syngens. De plus, au sein de chaque syngen, il existe un certain nombre de types d’accouplement généralement au nombre de deux.
Les types d’accouplement restent morphologiquement identiques mais ils présentent des différences physiologiques. Chez P. aurelia, il y a 14 syngènes et 28 types d’accouplement, alors que chez P. caudatum, il y a 16 syngènes et 32 types d’accouplement. On a observé qu’habituellement les paramécies ne se conjuguent ni avec les membres de leur propre type d’accouplement ni avec les autres variétés, mais seulement avec le deuxième type d’accouplement de leur propre variété.

Facteurs induisant la conjugaison :
Les facteurs induisant la conjugaison varient d’une espèce à l’autre mais certains d’entre eux sont donnés ci-dessous :
1. La conjugaison se produit généralement dans des conditions de vie défavorables ; la famine ou le manque de nourriture et un régime bactérien particulier ou certains produits chimiques sont réputés induire le processus de conjugaison chez certaines espèces de Paramecium.
2) La conjugaison se produit après environ 300 générations asexuées de fission binaire, ou bien elle alterne avec la fission binaire à de longs intervalles pour rajeunir le clone mourant, c’est-à-dire qu’elle se produit chez les individus qui doivent avoir traversé un nombre souhaitable de générations asexuées, dit période d’immaturité, puis ils deviennent sexuellement matures pour se conjuguer.
3 La conjugaison se produit lorsqu’il y a un changement dans la condition physiologique des paramécies, alors elle se produit entre ces individus qui sont un peu plus petits en taille (210 microns de long) et ils sont à un stade qui peut être considéré comme une période de vieillesse malsaine ; les paramécies de cette condition mourront si on ne leur permet pas de se conjuguer.
4. L’obscurité soudaine dans les conditions de lumière et les basses températures sont censées induire le processus de conjugaison chez certaines espèces.
5. La conjugaison n’a pas lieu pendant la nuit ou l’obscurité ; elle commence tôt le matin et se poursuit jusqu’à l’après-midi.
6. On dit qu’une substance protéique présente dans les cils des individus du type à accouplement induit la conjugaison.
Processus de conjugaison :
Le processus de conjugaison diffère dans les différentes espèces de Paramecium, mais le compte rendu sous-mentionné concerne le processus de conjugaison de P. caudatum (Fig. 20.21).
Dans la conjugaison, deux Paramecium caudatum (appelés pré-conjugants) de types d’accouplement opposés de la même variété se rapprochent par leurs surfaces ventrales et s’unissent par leurs rainures orales ; leurs cils produisent une substance à la surface du corps qui provoque l’adhésion des deux paramécies conjuguées.
Ils cessent de se nourrir et l’appareil de leur sillon oral disparaît. La pellicule et l’ectoplasme, au point de contact, des deux se brisent, et un pont protoplasmique se forme entre les deux animaux. Or, ces individus sont appelés conjugants.
Dans cet état, la paire de conjugants nage activement et, simultanément, une série de changements nucléaires ont lieu dans chaque conjugant, comme décrit ci-dessous :
Le macronoyau commence à se désintégrer, il devient de texture lâche et forme un écheveau complexe torsadé, pendant la dernière moitié de la période de conjugaison, il finira par disparaître en étant absorbé par le cytoplasme. Le micronoyau de chaque conjugué se divise deux fois, l’une d’elles étant une division réductrice.
Ainsi, quatre micronoyaux filles haploïdes sont produits dans chaque conjugant. Trois de ces quatre micronoyaux dégénèrent dans chacun, de sorte qu’il n’en reste qu’un seul.
Le micronoyau restant de chaque conjugant se divise mitotiquement en deux pronucléus ou noyaux gamétiques inégaux formant un pro-noyau femelle stationnaire plus grand et un pro-noyau mâle migrateur actif plus petit.
Le pro-noyau migrateur d’un conjugant traverse le pont protoplasmique et fusionne avec le pro-noyau stationnaire de l’autre conjugant pour former un synkaryon ou noyau de conjugaison dans lequel le nombre diploïde de chromosomes est restauré et il y a eu un échange de matériel héréditaire.
Le processus a été comparé à la fécondation chez les animaux supérieurs, mais il ne s’agit pas d’une fécondation car aucun gamète n’est impliqué. Les conjugants se séparent maintenant (après environ 12-48 heures) et sont appelés ex-conjugants. Le synkaryon de chaque ex-conjugant se divise trois fois pour former huit micronoyaux dans chaque ex-conjugant.
Quatre des huit micronoyaux s’agrandissent et deviennent des macronoyaux, et trois des quatre autres micronoyaux disparaissent.
Le micronoyau restant se divise et en même temps l’ex-conjugant se divise par fission binaire en deux cellules, chacune ayant deux macronoyaux et un micronoyau. Les cellules et leurs micronoyaux se divisent une seconde fois pour former quatre paramécies à partir de chaque ex-conjugant, de sorte que chacune possède un macronoyau et un micronoyau.
Le nouveau macronoyau, comme aussi le micronoyau, ont été constitués de matériel nouveau. Ces nouveaux noyaux contiennent probablement un potentiel nouveau et différent qui se reflète chez les individus sains.
Signification de la conjugaison :
Un clone s’éteindra si la réorganisation nucléaire ne se produit pas, mais le clone peut être rajeuni pour retrouver sa vigueur antérieure par un réarrangement nucléaire, cette réorganisation nucléaire est provoquée par la conjugaison, ainsi, la conjugaison est essentielle pour la poursuite de la fission binaire.
La signification de la conjugaison a été résumée ci-dessous :
1. La conjugaison sert de processus de rajeunissement et de réorganisation par lequel la vitalité de la race est restaurée. Si la conjugaison ne se produit pas pendant de longues périodes, les paramécies s’affaiblissent et meurent. (L’affirmation de Woodruffs de garder les paramécies en bonne santé pendant 22 000 générations sans conjugaison est réfutée par Sonneborn, car il a montré que toutes les paramécies de Woodruffs appartenaient au même type d’accouplement).
2. Il n’y a pas de distinction de sexe chez les conjugants bien que seuls les paramecia de deux types d’accouplement différents de la même variété se conjuguent.
3. Il n’y a pas de distinction de sexe, pourtant le pro-nucléus migrateur actif est considéré comme mâle et le pro-nucléus stationnaire comme la femelle.
4 La conjugaison n’est qu’une union temporaire, il n’y a pas de fusion du cytoplasme et aucun zygote n’est produit, mais le noyau de chaque ex-conjugant contient le matériel héréditaire de deux individus conjugués.
5. La conjugaison entraîne le remplacement du macronoyau par du matériel provenant du synkaryon, c’est un événement d’une importance fondamentale. Dans la fission binaire, les chromosomes du macronoyau étaient distribués au hasard dans les cellules filles, la poursuite de la fission binaire avait rendu le clone faible avec quelques anomalies structurelles.
La conjugaison entraîne la formation du nombre correct de chromosomes dans le macronucléus, de sorte que la race se renouvelle en vigueur. Le rôle du micronoyau est de rétablir un complexe chromosomique et génétique équilibré.
Comportement aberrant dans la reproduction chez Paramecium Caudatum :
Paramecium Caudatum montre certaines variations dans son comportement nucléaire pendant la fission et la conjugaison, ces déviations sont l’endomixie, l’autogamie, la cytogamie et l’hémixie. Dans les trois premiers processus, une recombinaison génétique est effectuée et un nouveau macronoyau est formé à partir du micronoyau.
(i) Endomixie :
Woodruff et Erdmann (1914) ont tout d’abord signalé un nouveau processus de réorganisation nucléaire, l’endomixie (Gr., endon = à l’intérieur ; mixis = mélange) chez Paramecium aurelia, une espèce bimicronucléée (Fig. 20.22). Ce processus a été décrit comme se produisant périodiquement et au cours duquel un nouvel appareil macro-nucléaire est produit sans formation de synkaryon. L’endomixie se produit au sein d’un seul individu.
Selon Woodruff et Erdmann, le macronoyau dégénère et les micronoyaux se divisent deux fois pour former huit micronoyaux. Six micronoyaux dégénèrent et deux restent. Comme il ne reste que deux micronoyaux, l’animal se divise par fission en deux cellules, chacune avec un micronoyau.
Le micronoyau de chaque cellule fille se divise deux fois pour former quatre micronoyaux. Deux micronuclei s’agrandissent pour former des macronuclei. L’animal et ses micronoyaux se divisent de façon à former deux individus filles, chacun ayant un macronoyau et deux micronoyaux.
L’endomixie se produit dans la variété de P. aurelia qui ne se conjugue pas, par conséquent, l’effet de l’endomixie peut être le même que celui de la conjugaison puisque les deux processus entraînent le remplacement du macronoyau par du matériel provenant du micronoyau, et les deux processus rajeunissent la vitalité de la race.
Mais les deux processus diffèrent car il n’y a pas de fusion des pronucléus dans l’endomixie ; l’endomixie peut être comparée à la parthénogenèse.
Cependant, certains travailleurs ont affirmé avec de bonnes raisons que l’endomixie n’est pas un processus valable et qu’elle a été décrite à cause d’une observation défectueuse. Selon toute probabilité, l’endomixie n’a pas lieu et il se peut qu’elle ne soit qu’un cas spécialisé d’autogamie.

Plus tard, Erdmamm et Woodruff (1916) ont rapporté une endomixie chez Paramecium caudatum. Diller, cependant, ne croit pas à la validité de ce processus et estime qu’Erdmann et Woodruff ont simplement combiné les étapes d’hémixie et d’autogamie en un seul schéma, l’endomixie.
(ii) Autogamie :
Diller (1934, 1936) et Sonneborn (1950) ont décrit un processus d’autofécondation ou autogamie se produisant chez un seul individu chez Paramecium aurelia (Fig. 20.23). Il a rapporté que dans l’autogamie, trois divisions micro-nucléaires (prégamiques), impliquant une maturation, produisent les noyaux gamétiques (pronucléi).
Durant l’autogamie chez P. aurelia, les deux micronoyaux se divisent deux fois (une fois méiotiquement) pour former huit micronoyaux, dont six dégénèrent. Pendant ce temps, le macronoyau se développe en une masse ressemblant à un écheveau qui se brise en morceaux pour être ensuite absorbé par le cytoplasme. Deux des huit micronoyaux, en tant que pronucléus, entrent dans un cône protoplasmique bombé près de la bouche de la cellule.
Les deux pronuclei fusionnent pour former un synkaryon. Le synkaryon se divise deux fois pour former quatre micronucléi. Deux micronuclei deviennent des macronuclei. La paramécie et ses micronoyaux se divisent pour former deux individus filles, chacun avec un macronoyau et deux micronoyaux. Ce processus s’achève en deux jours environ.
L’autogamie entraîne le rajeunissement de la race.
Elle ressemble à la conjugaison dans la mesure où le nouveau macronoyau est formé par du matériel provenant du micronoyau, dans le nouveau macronoyau le nombre correct de chromosomes est rétabli ; et aussi dans le fait que la fusion de deux pronucléus se produit. Mais l’autogamie diffère de la conjugaison car un seul individu participe à l’autogamie et il fournit les deux pronuclei, c’est une sorte d’auto-fécondation.
(iii) L’hémixie :
Diller (1936) a signalé l’hémixie chez Paramecium aurelia (figure 20.24). L’hémixie est principalement un processus de fragmentation et de division macro-nucléaire sans activité micro-nucléaire inhabituelle. Diller a classé l’hémixie en quatre types, à savoir A, B, C et D comme le montre la Fig. 20.24 chez P. aurelia mais il a également rencontré tous les types dans les cultures de masse de P. caudatum et P. multimicronucleatum.
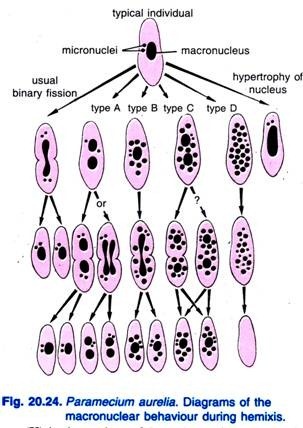
Le type A est la forme la plus simple d’hémixie caractérisée par une division du macronoyau en deux parties ou plus. Cette division n’est pas synchronisée avec la division du micro-noyau.
Le type B est caractérisé par l’extrusion d’une à 20 ou plusieurs boules de chromatine du macronoyau dans le cytoplasme.
Le type C est caractérisé par la division simultanée du macronoyau en deux ou plusieurs portions majeures et l’extrusion de boules macro-nucléaires dans le cytoplasme.
Le type D est considéré comme représentant des conditions pathologiques dans lesquelles le macronoyau subit une fragmentation complète en boules de chromatine qui finissent par disparaître de la cellule. Les micronoyaux disparaissent généralement avant la dissolution du macronoyau.
(iv) La cytogamie :
Wichterman (1939) a signalé un autre processus sexuel chez Paramecium caudatum, qu’il a appelé cytogamie. Dans la cytogamie, il n’y a pas d’échange nucléaire. Dans ce processus, deux individus se rapprochent par leurs surfaces ventrales, mais la pellicule des deux individus ne se rompt pas.
Le micronoyau de chaque individu se divise trois fois pour former huit micronoyaux, dont six se désintègrent dans chaque individu. Les deux micronoyaux restants fusionnent pour former un synkaryon dans chaque cellule. Les animaux se séparent maintenant.
La cytogamie diffère de l’autogamie dans la mesure où il y a deux animaux en contact l’un avec l’autre, mais elle ressemble à l’autogamie et à la conjugaison dans la fusion de deux pronuclei. La cytogamie diffère de la conjugaison en ce qu’il n’y a pas d’échange nucléaire entre les deux animaux qui se rapprochent.
Certaines particules cytoplasmiques signalées dans le Paramecium Caudatum :
(i) Particules kappa :
En 1938, T.M. Sonneborn a rapporté que certaines races (connues sous le nom de tueurs ou souche tueuse) de Paramecium produisent une substance toxique, appelée paramécine qui est mortelle pour d’autres individus appelés sensibles. La paramécine est soluble dans l’eau, diffusible et dépend pour sa production de certaines particules situées dans le cytoplasme de la Paramécie (souche tueuse).
Ces particules sont appelées particules kappa. Les particules kappa contiennent de l’ADN et de l’ARN. Une Paramecium tueuse peut contenir des centaines de particules kappa. L’étude détaillée de ces particules a révélé qu’un gène dominant (K) dans le noyau de la Paramecium est nécessaire pour que les particules kappa existent, se multiplient et produisent de la paramécine.
(ii) mµ, Particules :
R.W. Siegel (1952) a signalé un autre type de particules tueuses dans le cytoplasme de certaines Paramecium. Une paramécie avec des particules mp est appelée mate killer car lorsqu’elle se conjugue avec une paramécie sans particules mµ appelée mate sensitive, alors elle tue cette dernière. Les particules mp sont également composées d’ADN, d’ARN, etc.
Ces particules n’existent que chez les paramécies dont le micronoyau contient au moins un gène dominant de l’une ou l’autre des deux paires de gènes chromosomiques non liés (M1 et M2).
(iii) Particules Pi :
Ces particules sont censées être la forme mutante des particules kappa mais elles ne produisent aucun type de substance toxique.
(iv) Particules Lambda :
Ces particules sont signalées dans les paramécies tueuses et sont censées produire une certaine substance responsable de la lyse ou de la désintégration des paramécies sensibles, c’est-à-dire qui ne la possèdent pas.