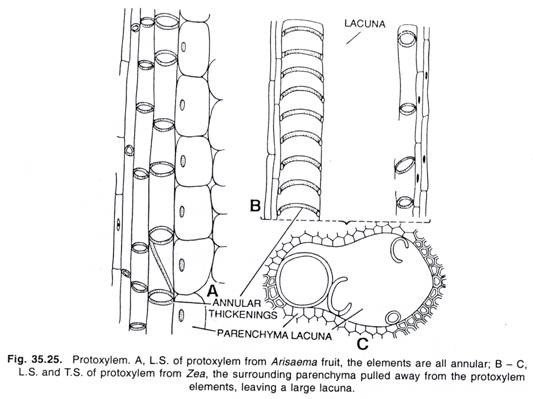
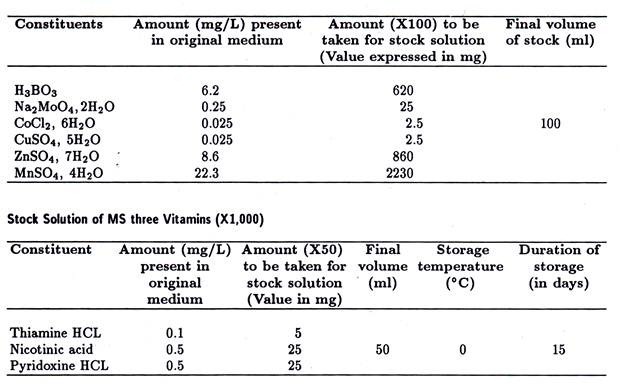
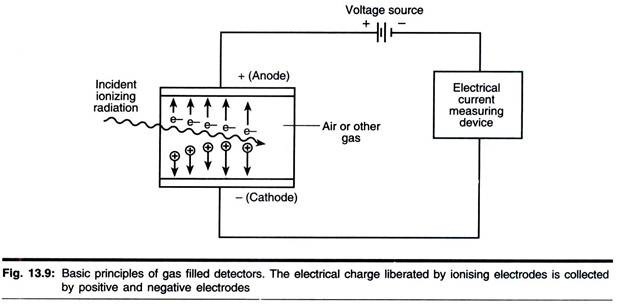
ADVERTISEMENTS:
En este artículo hablaremos del Paramecium Caudatum:- 1. Hábito, Hábitat y Cultivo del Paramecium Caudatum 2. Estructura del Paramecium Caudatum Estructura de Paramecium Caudatum 3. Locomoción 4. Nutrición 5. Respiración y excreción 6. Osmorregulación 7. Comportamiento 8. Reproducción 9. Comportamiento aberrante en la reproducción 10. Algunas partículas citoplasmáticas.
Contenido:
- Hábitat, Hábitat y cultivo de Paramecium Caudatum
- Estructura de Paramecium Caudatum
- Locomoción de Paramecium Caudatum
- Nutrición en Paramecium Caudatum
- Respiración y Excreción en Paramecium Caudatum
- Osmorregulación de Paramecium Caudatum
- Comportamiento de Paramecium Caudatum
- Reproducción en Paramecium Caudatum
- Comportamiento aberrante en la reproducción en Paramecium Caudatum
- Algunas partículas citoplasmáticas reportadas en Paramecium Caudatum
- Hábitat, Hábitat y cultivo de Paramecium Caudatum:
- Cultivo de Paramecium:
- Estructura de Paramecium Caudatum:
- (i) Tamaño y forma de Paramecium Caudatum:
- (ii) Pellicula:
- (iii) Cilios:
- (iv) Sistema infraciliar:
- (v) Ranura oral y citopigia:
- (vi) Citoplasma:
- (vii) Ectoplasma:
- (viii) Triquistes:
- (ix) Neuromotorium and Associated Fibrils:
- (x) Endoplasma:
- (xi) Vacuolas contráctiles:
- (xii) Vacuolas alimenticias:
- Locomoción de Paramecium Caudatum:
- (i) Metabolia o contorsiones del cuerpo:
- (ii) Locomoción ciliar:
- Nutrición del Paramecium Caudatum:
- Mecanismo de alimentación:
- Digestión y Egestión del Paramecium Caudatum:
- Respiración y Excreción de Paramecium Caudatum:
- Comportamiento de Paramecium Caudatum:
- (i) Reacciones al contacto (Thigmotaxis):
- (ii) Reacciones a productos químicos (Quimiotaxis):
- (iii) Reacciones a la temperatura (Termotaxis):
- (iv) Reacciones a la luz (Fototaxis):
- (v) Reacciones a la corriente eléctrica (Galvanotaxis):
- (vi) Reacciones a la corriente de agua (reotaxis):
- (vii) Reacciones a la gravedad (Geotaxis):
- Reproducción en Paramecium Caudatam:
- (i) Fisión binaria transversal:
- (ii) Conjugación:
- Comportamiento aberrante en la reproducción en Paramecium Caudatum:
- (i) Endomixis:
- (ii) Autogamia:
- (iii) Hemixis:
- (iv) Citogamia:
- Algunas partículas citoplasmáticas reportadas en Paramecium Caudatum:
- (i) Partículas Kappa:
- (ii) mµ, partículas:
- (iii) Partículas Pi:
- (iv) Partículas Lambda:
Hábitat, Hábitat y cultivo de Paramecium Caudatum:
Paramecium caudatum (Gr., paramekes = oblongo; L., caudata = cola) se encuentra comúnmente en estanques de agua dulce, piscinas, zanjas, arroyos, lagos, embalses y ríos. Se encuentra especialmente en abundancia en estanques estancados ricos en materia en descomposición, en infusiones orgánicas y en las aguas residuales. Paramecium caudatum es un organismo de vida libre y su distribución es mundial.
Cultivo de Paramecium:
ADVERTENCIAS:
Toma hierbas sumergidas de un estanque y colócalas en una jarra con agua destilada, tapa la jarra y deja que se pudra; en pocos días aparecerán enjambres de Paramecia. Ahora hervir heno en agua, decantar la infusión y añadir unos granos de trigo, y dejarla reposar hasta que esté turbia de bacterias.
Transfiera los Paramecia del primer frasco a este líquido donde se multiplicarán rápidamente. Las infusiones de heno por sí solas producirán Paramecia mostrando la presencia de quistes, y se ha informado de quistes parecidos a granos de arena, pero no hay pruebas de que los Paramecia formen quistes, ya que nunca se han confirmado.
Estructura de Paramecium Caudatum:
(i) Tamaño y forma de Paramecium Caudatum:
Paramecium caudatum (Fig. 20.1) es un organismo microscópico y visible a simple vista como un diminuto cuerpo alargado.
ADVERTENCIAS:
Se presenta de color gris claro o blanco y mide comúnmente entre 170 y 290 micras de longitud y puede alcanzar una longitud de hasta 300-350 micras. P. caudatum se parece a la suela de una zapatilla o zapato, de ahí que el animal se conozca comúnmente como animalcule de zapatilla. Es cuatro veces más largo que ancho y algo cilíndrico con extremos claramente diferenciados.
La parte anterior, que se desplaza hacia delante, es delgada con un extremo romo o redondeado, mientras que el extremo posterior es algo puntiagudo o cónico. La parte más ancha del organismo está justo por debajo de la mitad. El cuerpo del animal es de forma asimétrica mostrando una superficie oral o ventral bien definida y una aboral o dorsal.
(ii) Pellicula:
El cuerpo está cubierto por una fina película de doble capa, elástica y firme, hecha de gelatina. La película mantiene la forma del animal, pero es lo suficientemente elástica para permitir las contracciones. La película tiene doble membrana, la exterior es continua con los cilios y la interior con el ectoplasma. Bajo el mayor aumento del microscopio, la película muestra depresiones rectangulares o hexagonales en su superficie.

Esta disposición se conserva en la superficie dorsal del Paramecium pero en la superficie ventral las crestas convergen por delante y por detrás hacia una apertura preoral y postoral. Cada depresión hexagonal está perforada por una abertura central por la que sale un solo cilio. Los márgenes anterior y posterior de las depresiones hexagonales llevan las aberturas de los tricocistidios.
El estudio microscópico electrónico de la película (Fig. 20.2) realizado por Ehret y Powers (1957) ha revelado que las depresiones hexagonales corresponden a series regulares de cavidades, los alvéolos. Todos los alvéolos forman colectivamente una capa alveolar continua, que está delimitada por una membrana alveolar externa y otra interna.
La capa externa se encuentra en estrecho contacto bajo la membrana celular externa. Por lo tanto, la película incluye la membrana celular externa, la membrana alveolar externa y la membrana alveolar interna.
(iii) Cilios:
Todo el cuerpo está cubierto de numerosas y pequeñas proyecciones similares a pelos llamadas cilios. Los cilios se presentan en hileras longitudinales por todo el cuerpo, esta condición se conoce como holotricos en la que los cilios del cuerpo son iguales. Los cilios tienen la misma estructura que los flagelos, tienen una vaina protoplásmica externa o membrana plasmática con nueve fibrillas longitudinales dobles en un anillo periférico. En algunos cilios las nueve fibrillas exteriores no están emparejadas.
ADVERTENCIAS:
Hay dos fibrillas longitudinales centrales que son más delgadas que las externas. Cada cilio surge de un gránulo basal o cinetosoma. Los nueve pares de fibrillas periféricas se fusionan para formar la pared del cinetosoma, por lo tanto, el cinetosoma es un tubo que está abierto o cerrado en su extremo inferior, las dos fibrillas centrales se detienen a nivel de la película en la mayoría de los ciliados.
Del cinetosoma sale un fino rizoplasto que no se une al núcleo. Muchos Metazoos también tienen cilios, su estructura es la misma, excepto que el gránulo basal es diferente y tiene finos filamentos o fibras de enraizamiento que se extienden hasta el citoplasma. Pero los cilios se diferencian de los flagelos por ser generalmente más numerosos y de menor tamaño.
La ciliatura puede dividirse convenientemente en cilios corporales o somáticos, que se encuentran en la superficie del cuerpo, y en ciliatura oral, que está asociada a la región bucal. Los cilios corporales son iguales pero son más largos en el extremo posterior, de ahí el nombre de caudatum. Los cilios son orgánulos de locomoción y recolección de alimento, también actúan como receptores sensoriales y detectan los estímulos del medio externo.

Ultraestructura de los cilios:
Los cilios y flagelos tienen una composición fibrilar. En la base, el cilio tiene un diámetro de aproximadamente 0,2 micras o 2.000 A0 que puede estar hasta 10 micras por encima de la superficie celular. Los cilios están limitados por una membrana unitaria de 90 A0 de grosor que se asemeja a la membrana plasmática y permanece continua con ella. El espacio delimitado del cilio contiene una sustancia acuosa conocida como matriz.
En la matriz quedan incrustadas once fibrillas longitudinales o microtúbulos. De las once fibrillas, dos están situadas en el centro, mientras que las nueve restantes permanecen dispuestas periféricamente alrededor de las fibrillas centrales. Cada una de las nueve fibrillas exteriores tiene un diámetro de 360A0 y está compuesta por dos subfibrillas de 180 a 250A0 de diámetro.
Estas subfibrillas se designan como la subfibrilla A y la subfibrilla B.
ADVERTENCIAS:
La subfibrilla A es ligeramente más grande que la subfibrilla B. La subfibrilla A emite dos gruesas proyecciones o brazos por un lado. Los brazos de la subfibrilla A de todas las fibrillas exteriores permanecen dirigidos en el sentido de las agujas del reloj. Además, la subfibrilla A está más cerca del centro del cilio que la subfibrilla B. Ambas subfibrillas tienen una pared común de 50A0 de grosor.
Las dos fibrillas centrales no tienen subfibrillas emparejadas como las nueve fibrillas periféricas, sino que cada una contiene un solo túbulo. Cada fibrilla central tiene un diámetro de unos 250A0 y está compuesta por una pared de 60A0 de grosor.
Las dos fibrillas centrales permanecen separadas por un espacio de 350A0 y quedan envueltas en una vaina común. Gibbnos (1967) ha informado de que la vaina de las fibrillas centrales da salida a nueve enlaces o radios orientados radialmente a cada subfibrilla A.

La microscopía electrónica de alta resolución ha revelado que cada una de las fibrillas periféricas y centrales de los cilios y flagelos está compuesta por diez a doce filamentos de 40A0 de grosor. Cada filamento está provisto de cuentas. Cada cuenta permanece dispuesta en el entramado de 40 por 50A0 en el plano de la pared del túbulo. Estas perlas se consideran como la subunidad básica de la estructura del túbulo.
(iv) Sistema infraciliar:
El sistema infraciliar está situado justo debajo de los alvéolos peliculares. Está formado por el cinetosoma o cuerpo basal y el cinetodesma. Los cilios surgen de los cinetosomas y de cada cinetosoma surge una delicada fibrilla citoplasmática llamada cinetodesma, (Fig. 20.2).
Debajo de la película, ligeramente a la derecha, pero unido a todos los cinetosomas de una fila longitudinal, se encuentra un haz longitudinal de varios cinetodesmos, un cinetodesmo de cada cinetosoma se extiende por una distancia anterior en su propio haz de cinetodesmos.
Una hilera longitudinal de cinetosomas con sus cinetodesmos forma una unidad longitudinal llamada cineto. Todos los cinetosomas o cinetosomas forman el sistema infraciliar de un ciliado. Los cinetos se encuentran en la corteza por debajo de la película, su número es bastante constante para cada ciliado.
El sistema infraciliar controla y coordina los movimientos de los cilios, y provoca la formación de orgánulos en la división celular, por ejemplo, algunos cinetos forman la boca. En la fisión binaria de los ciliados los cinetos se cortan transversalmente en dos, cada uno de los cuales va a una célula hija, lo que se denomina fisión perikinetal.
(v) Ranura oral y citopigia:
En el lado ventrolateral hay una gran depresión oblicua y poco profunda llamada surco oral o peristoma que da al animal un aspecto asimétrico. Corre oblicuamente hacia atrás desde un lado (generalmente de izquierda a derecha pero en algunos casos de derecha a izquierda) y termina un poco detrás del cuerpo medio. El surco oral desemboca en una corta depresión cónica en forma de embudo llamada vestíbulo.
ADVERTENCIAS:
El vestíbulo desemboca directamente en la abertura fija de forma ovalada llamada citostoma (boca). Desde el citostoma se extiende directamente hacia el centro del cuerpo la amplia citofaringe. La citofaringe gira entonces bruscamente hacia la parte posterior para convertirse en el esófago, que es delgado y se estrecha.
Así, el esófago es aproximadamente paralelo a la superficie del cuerpo del Paramecium excepto en su extremo posterior. Aquí el esófago vuelve a girar hacia el centro del animal para desembocar en la vacuola alimenticia en formación.
El citopigio (también llamado ano celular o mancha anal o citoprocto) se encuentra en la superficie ventral del cuerpo casi verticalmente detrás del citostoma o boca. Las partículas de alimento no digeridas se eliminan a través del citopígono. La ciliación de la citofaringe es muy complicada.
Gelei (1934) informó de la presencia de cuatro filas y Lund (1941) observó al menos cuatro filas. En la pared izquierda de la citofaringe se encuentra una estructura llamada pennícula, que gira en espiral aproximadamente 90 grados, de modo que su extremo posterior se encuentra en la superficie oral (ventral) del esófago.
Según Lund, el penículo consta de ocho hileras de cilios dispuestas en dos bloques estrechamente dispuestos de cada uno. Una banda similar compuesta por cuatro filas de cilios largos, menos compactos que en el penículo, se denomina cuádruple. Desciende en espiral por la pared dorsal de la cavidad bucal y termina cerca del penículo.
El penniculus y el quadrulus han sido llamados erróneamente una membrana ondulante por algunos trabajadores. El cuadrilátero y el penículo controlan el paso del alimento. No se sabe cómo funcionan los cilios, probablemente sus fibrillas se contraen de forma rítmica lo que provoca su flexión. Gelei (1925) señaló que la función del penículo es forzar el paso de los elementos alimenticios al interior del organismo.

(vi) Citoplasma:
El citoplasma se diferencia en una zona estrecha, externa o cortical, llamada ectoplasma y una región mayor, interna o medular, llamada endoplasma.
(vii) Ectoplasma:
El ectoplasma (ectosarc o corteza) es una parte permanente del cuerpo, llamativamente delimitada del endoplasma. El ectoplasma forma una capa exterior firme, clara, fina y densa. Contiene los triquistes, los cilios y las estructuras fibrilares y está delimitado externamente por una cubierta llamada película.
(viii) Triquistes:
Incorporados en el ectoplasma, en ángulo recto con la superficie, se encuentran pequeños sacos fusiformes llamados tricocistos. Un pequeño punto en cada margen anterior y posterior del hexágono marca la posición de un tricocisto. Están llenos de un líquido refractario y denso que tiene una sustancia hinchante, en el extremo exterior hay una cabeza cónica o espiga.
Los tricocistos se encuentran perpendiculares en el ectoplasma, se abren por pequeños poros en las crestas de las zonas hexagonales de la película.
Surgen de los cinetosomas de los cilios, luego migran y se sitúan a igual distancia en el endoplasma. Cuando el animal se irrita, los tricocistos se descargan como largos hilos pegajosos. Un tricocisto descargado tiene una espiga opaca, como una uña invertida, y un largo eje estriado, pero el eje no se ve en el estado no descargado y probablemente se forma durante la descarga.
La función de los triquistes es incierta, pero se descargan como reacción a los contactos locales y a las lesiones, pueden servir como orgánulos de defensa.
Pero esto es incierto porque los tricocistos son ineficaces contra Didinium, el principal depredador de Paramecium, pueden servir para fijar al animal a un punto durante la alimentación. En algunos ciliados, los tricocistos actúan como orgánulos de ofensa. Después de la descarga de los triquistes, se regeneran a partir de los cinetosomas.


(ix) Neuromotorium and Associated Fibrils:
Según Lund (1933) en la pared dorsal izquierda de la citofaringe, aproximadamente al nivel del margen posterior del citostoma, se encuentra una masa muy pequeña y bilobulada, el neuromotorium. Desde el neuromotorio, las fibrillas irradian hacia el endoplasma.
De ellas, cuatro o más suelen pasar casi hasta la pared dorsal del cuerpo, pero el resto son más cortas y no tienen una posición definida. Todas se denominan fibrillas endoplásmicas. Su función es desconocida, pero pueden coordinar los movimientos de alimentación de los cilios orales. Las fibrillas también pueden dar soporte mecánico, elasticidad, contractilidad, conductividad e influencia metabólica.

(x) Endoplasma:
El endoplasma o médula es la parte más fluida y voluminosa del citoplasma que contiene muchos gránulos citoplasmáticos así como otras inclusiones y estructuras de naturaleza especializada. Las inclusiones citoplasmáticas son mitocondrias, aparatos de Golgi, vacuolas, cristales, gránulos y cromidios, etc. También se encuentran en el endoplasma otras estructuras, a saber, núcleos, vacuolas contráctiles y vacuolas alimenticias.
Núcleos:
En el endoplasma cerca del citostoma se encuentran dos núcleos, es decir, el Paramecium es heterocariótico, un gran macronúcleo elipsoidal y granular y otro pequeño micronúcleo compacto.
El macronúcleo es un cuerpo conspicuo, elipsoidal o con forma de riñón. Es de tipo compacto y contiene hilos finos y gránulos discretos de cromatina muy apretados, de tamaño variable e incrustados en una matriz acromática. Posee muchos nucléolos y mucho más material de cromatina (ADN).
Es un núcleo somático o vegetativo. Se divide amitóticamente y controla las funciones vegetativas (actividades metabólicas) del animal. Se somete a la mitosis.
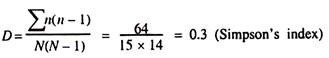
El micronúcleo es pequeño, compacto y esférico. Se encuentra generalmente cerca del macronúcleo, a menudo en una concavidad. Los gránulos e hilos finos de cromatina están distribuidos uniformemente por toda la estructura. El micronúcleo se divide mitóticamente y controla la reproducción.
El número de micronúcleos varía con la especie; es uno en P. caudatum, dos en P. aurelia y muchos en P. multimicronucleatum. El micronúcleo contiene un nucleolo distinto en P. aurelia, pero no se encuentra en P. caudatum. Moses (1949; 1950) ha informado de que el macronúcleo y el micronúcleo son idénticos en su composición química.
(xi) Vacuolas contráctiles:
Hay dos grandes vacuolas contráctiles llenas de líquido, cada una situada cerca de un extremo del cuerpo, cerca de la superficie dorsal. Su posición es fija (a diferencia de la Ameba), se encuentran entre el ectoplasma y el endoplasma, pero son orgánulos temporales que desaparecen periódicamente. En algunas especies, parecen tener una membrana de revestimiento, en cuyo caso no desaparecen por completo durante la sístole.
Conectados a cada vacuola contráctil hay de cinco a doce canales tubulares radiantes, cada uno de los cuales consta de una parte terminal, una ampolla larga que se colapsa cuando está vacía, y un canal inyector corto que se abre en la vacuola.

Los canales comunican con una gran parte del cuerpo de donde toman líquidos y los vierten en la vacuola que, de este modo, se reconstituye y aumenta de tamaño, cuando la vacuola contráctil alcanza su tamaño máximo se contrae bruscamente (sístole) y descarga su contenido a través de un poro permanente en la película, entonces los canales vuelven a formar las vacuolas contráctiles, los canales no desaparecen del todo ya que son estructuras permanentes.
Las dos vacuolas contráctiles descargan irregularmente, la posterior se contrae más rápidamente porque está cerca de la citofaringe y entra más agua en ella. La función principal de los canales y las vacuolas contráctiles es hidrostática, eliminan el exceso de agua del protoplasma, el agua es en parte absorbida y en parte tomada durante la alimentación.
La sustancia de desecho nitrogenada tiene compuestos de amoníaco y algunos uratos que son expulsados de las vacuolas contráctiles junto con el CO2 pero no hay evidencia de que la materia excretora sea secretada por el protoplasma en los canales o vacuolas contráctiles.
Hay un citoplasma o ano permanente fijo debajo del peristoma a un lado de la citofaringe, a través del cual se expulsan los restos no digeridos de alimentos y los uratos.
El estudio microscópico electrónico de las vacuolas contráctiles ha revelado que cada aparato contráctil está formado por algunos de los túbulos del retículo endoplásmico, túbulos nefridiales, canales alimentadores, vacuolas accesorias (canales radiales) y vacuola contráctil principal. Se supone que las vacuolas accesorias son las ampollas de los canales alimentadores (Fig. 20.11).
(xii) Vacuolas alimenticias:
Son cuerpos aproximadamente esféricos, no contráctiles, que varían en tamaño y número y que se encuentran en el endoplasma. Contienen partículas alimenticias ingeridas, principalmente bacterias, y una pequeña cantidad de líquido delimitado por una fina membrana definida. Volkonsky (1934) propuso el nombre de gastriolas para estas vacuolas. Asociadas a las vacuolas alimentarias están los gránulos digestivos.
Locomoción de Paramecium Caudatum:
Paramecium Caudatum realiza la locomoción por dos métodos, a saber, metabolismo o contorsiones del cuerpo y por cilios.
(i) Metabolia o contorsiones del cuerpo:
El cuerpo del Paramecium Caudatum posee elasticidad, puede apretarse a través de un pasaje más estrecho que su cuerpo, después de lo cual el cuerpo asume su forma normal. Este cambio temporal de la forma del cuerpo es el metabolismo, se produce en el Paramecium por el protoplasma.
(ii) Locomoción ciliar:
La locomoción provocada por los cilios es el método principal. Los cilios pueden batir hacia delante o hacia atrás, lo que permite al animal nadar hacia delante o hacia atrás.
Normalmente el animal nada hacia delante, los cilios laten hacia atrás pero de forma oblicua, los cilios se ponen rígidos y se doblan hacia atrás rápidamente hasta casi tocar la superficie del cuerpo, esto se llama la brazada efectiva; luego los cilios se vuelven blandos y vuelven lentamente a la posición vertical original, esto se llama , brazada de recuperación.
Los cilios de la misma fila transversal laten juntos y los de la misma fila longitudinal laten uno tras otro desde el extremo anterior al posterior.

Este movimiento coordinado de los cilios se denomina ritmo metacronal, que se debe al sistema infraciliar; esto provoca la natación hacia delante por parte del animal. Pero cuando los cilios del cuerpo laten oblicuamente hacia atrás, entonces al mismo tiempo los cilios más largos del surco bucal laten más vigorosamente lo que hace que el extremo anterior se desvíe hacia la izquierda.
La acción de los cilios del cuerpo y del surco oral hace que el animal rote sobre su eje largo. Esta rotación es siempre hacia la izquierda (excepto en P. calkinsi que gira en espiral hacia la derecha).
Esta combinación de movimiento hacia delante, giro y rotación hace que el animal se mueva hacia delante en una trayectoria en espiral en sentido contrario a las agujas del reloj. Esta trayectoria tiene un eje recto, y la misma superficie corporal del animal permanece hacia el eje de la trayectoria en espiral. Pero al nadar hacia atrás, todas las especies giran hacia la derecha.
El latido ciliar puede invertirse de manera que los cilios se mueven oblicuamente hacia delante por lo que el animal nada hacia atrás. Por acción ciliar, el Paramecium se mueve con una velocidad de 1500 micras o incluso más por segundo.
Jennings sostuvo que el movimiento en espiral del Paramecium se debe a que, mientras los cilios golpean principalmente hacia atrás, lo hacen oblicuamente hacia la derecha, provocando así que el animal se vuelque hacia la izquierda.
También este giro del cuerpo hacia la superficie aboral se debe en gran medida a la mayor potencia del golpe efectivo de los cilios orales que golpean más directamente hacia atrás. El resultado -la rotación del Paramecium sobre su eje largo- permite al Paramecium seguir un curso más o menos recto al formar grandes espirales.
Nutrición del Paramecium Caudatum:
En el Paramecium Caudatum, la nutrición es holozoica. El alimento comprende principalmente bacterias y diminutos protozoos. El Paramecium no espera el alimento sino que lo caza activamente.
Se afirma que el Paramecium Caudatum muestra una elección en la selección de su alimento, pero no parece haber ninguna base para esto aunque engulle sólo ciertos tipos de bacterias; los datos disponibles sugieren que de 2 a 5 millones de individuos de Bacillus coli son devorados por un solo Paramecium en 24 horas. También se alimenta de plantas unicelulares como algas, diatomeas, etc., y de pequeños trozos de animales y vegetales.
Mecanismo de alimentación:
Cuando el Paramecium Caudatum entra en una región de alimento abundante, viene a descansar. Se alimenta sólo en reposo o cuando nada muy lentamente, nunca se alimenta cuando nada rápido. El batir de los cilios del surco oral hace que un vórtice en forma de cono de agua cargada de alimento sea barrido hacia el surco oral desde una distancia anterior al extremo anterior (Fig. 20.16).
Las partículas de alimento pasan entonces al vestíbulo desde donde algunas partículas de alimento son rechazadas y expulsadas, pero otras pasan al citostoma.
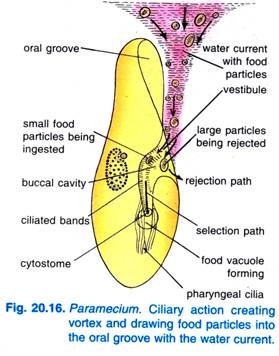
Al final de la citofaringe se forma una vacuola alimentaria que se llena de partículas de alimento. Los cuádriceps y los penículos controlan el paso del alimento a la vacuola alimentaria que se forma lateralmente. Cuando la vacuola alimenticia alcanza un cierto tamaño, las fibras postbucales sujetan la vacuola alimenticia y ésta es pinzada por ellas y comienza su curso.
La vacuola contiene algo de agua además del alimento. Los movimientos rotativos del endoplasma, llamados ciclosis, llevan a las vacuolas alimenticias a lo largo de un curso definido que es funcionalmente equivalente a un tracto digestivo.
El tracto comienza desde el final de la citofaringe, luego hacia la parte posterior, luego hacia delante para circular con el endoplasma, luego hacia la superficie dorsal, luego hacia el extremo anterior, luego hacia abajo hasta la citofaringe. Al principio de su recorrido la vacuola alimenticia disminuye de tamaño, luego vuelve a aumentar.

Digestión y Egestión del Paramecium Caudatum:
Durante la ciclosis, la digestión se produce por las enzimas secretadas por el protoplasma en las vacuolas. En la digestión, las proteínas se transforman en aminoácidos, los carbohidratos en azúcares solubles y glucógeno, y probablemente también se digieren las grasas.
El contenido de las vacuolas de los alimentos es al principio ácido (pH alrededor de 4) y luego se vuelve alcalino, la mayor digestión ocurre durante la fase alcalina. La materia no digerida es egestada a través del citopié con cierta fuerza.
La ciclosis puede demostrarse experimentalmente; si se alimenta al Paramecium con leche teñida con rojo Congo, los glóbulos de grasa de la leche en las vacuolas alimenticias se volverán primero rojos debido a la reacción ácida de las enzimas, luego cambiarán de tonos de púrpura a azul debido a la reacción alcalina, las vacuolas mostrarán el curso de la ciclosis.
Respiración y Excreción de Paramecium Caudatum:
El intercambio de gases (oxígeno y dióxido de carbono) tiene lugar a través de la película semipermeable como otros protozoos de agua dulce por el proceso de difusión. El Paramecium Caudatum obtiene su oxígeno del agua circundante. El dióxido de carbono y los desechos orgánicos, como el amoníaco, resultantes del metabolismo, son probablemente excretados por difusión hacia el agua en la dirección inversa.

6. Osmorregulación en Paramecium Caudatum:
Paramecium Caudatum tiene dos vacuolas contráctiles, una anterior y otra posterior. La función de las vacuolas contráctiles es la osmorregulación, es decir, regular el contenido de agua del cuerpo y puede servir también en la excreción de desechos nitrogenados como la urea y el amoníaco.
El exceso de agua (debido a la endosmosis continua) dentro del citoplasma se segrega en los túbulos del retículo endoplásmico y va a los túbulos nefridiales → canales alimentadores → y se recogen en la ampolla de una serie de 6 a 11 canales radiantes que convergen hacia y descargan en cada vacuola. Los canales son más conspicuos cuando se está formando una vacuola.
Cuando cada vacuola se hincha (diástole) hasta un cierto tamaño, se contrae (sístole) y descarga al exterior probablemente a través de un poro. Las vacuolas contráctiles se contraen alternativamente, a intervalos de 10 a 20 segundos.
La vacuola contráctil posterior trabaja más rápido que la anterior debido a la entrada de gran cantidad de agua en la región posterior por la citofaringe. Las vacuolas contráctiles mantienen una concentración óptima de agua en el citoplasma del cuerpo eliminando el exceso.

Comportamiento de Paramecium Caudatum:
Las respuestas de Paramecium Caudatum a diversos tipos de estímulos se aprenden mediante el estudio de sus reacciones y de la agrupación o dispersión de los individuos en un cultivo. La respuesta es positiva si el animal se mueve hacia un estímulo y negativa cuando se aleja. Ante un estímulo adverso el animal sigue dando la reacción de evitación hasta que escapa.
En la reacción de evitación, el latido ciliar se invierte, el animal se mueve hacia atrás una corta distancia y luego gira en una trayectoria cónica girando el extremo anterior aboralmente mientras pivota sobre el extremo posterior. Todos los ajustes se realizan por ensayo y error. Los experimentos han demostrado que el extremo anterior del animal es más sensible que las otras partes.
Las respuestas del Paramecium a los diferentes estímulos pueden agruparse como sigue:
(i) Reacciones al contacto (Thigmotaxis):
La respuesta al contacto es variada en el Paramecium. Si se toca ligeramente el extremo anterior con una punta fina, se produce una fuerte reacción de evitación. Cuando un Paramecium nadando choca con algún objeto en el agua, pero si se le toca en otro lugar puede no haber respuesta. Un individuo que se mueve lentamente suele responder positivamente al contacto con un objeto, posándose sobre él.
(ii) Reacciones a productos químicos (Quimiotaxis):
Generalmente los Paramecios responden a un estímulo químico mediante la evitación de la reacción. Si se introduce una gota de solución salina débil (0,5 por ciento) en una población de Paramecios en un microdeslizador, los animales responden con la reacción de evitación y ninguno entra en la gota. Para los ácidos, sin embargo, la respuesta es positiva incluso cuando la concentración es lo suficientemente fuerte como para matarlos.
(iii) Reacciones a la temperatura (Termotaxis):
El Paramecium busca una temperatura óptima de 24 a 28°C. Cuando se produce un cambio de temperatura muy por encima o por debajo del rango óptimo, los Paramecios muestran una reacción de evitación. Un mayor calor estimula el movimiento rápido y las reacciones de evasión hasta que los animales escapan o se matan.
(iv) Reacciones a la luz (Fototaxis):
Con la excepción del Paramecium bursaria verde, que es positivamente fototáctico, las demás especies son indiferentes a la luz ordinaria. Sin embargo, cuando la intensidad de la luz se incrementa repentina y bruscamente, generalmente se produce una reacción negativa. Los paramecios muestran una respuesta negativa inmediata a los rayos ultravioleta.
(v) Reacciones a la corriente eléctrica (Galvanotaxis):
Los paramecios responden a los estímulos eléctricos. Cuando se colocan dos electrodos opuestos en un plato poco profundo que contiene Paramecia y se aplica una corriente constante, todos los organismos nadan en la misma dirección hacia el cátodo o electrodo negativo donde se concentran en gran número.
Si se invierte la dirección de la corriente eléctrica mientras los Paramecia nadan hacia el cátodo, los organismos invierten la dirección y nadan hacia el nuevo cátodo.

(vi) Reacciones a la corriente de agua (reotaxis):
El Paramecio muestra una reotaxis positiva. En una corriente de agua suave los Paramecia se moverán mayoritariamente con la corriente con sus extremos anteriores río arriba.
(vii) Reacciones a la gravedad (Geotaxis):
Los Paramecia generalmente exhiben una respuesta negativa a la gravedad como se observa en un cultivo donde muchos individuos se reúnen cerca bajo la película superficial con sus extremos anteriores más arriba. Si se introducen los Paramecios en un tubo en forma de U invertido lleno de agua y tapado en ambos extremos, se desplazan inmediatamente hacia arriba en la parte horizontal del tubo.
Reproducción en Paramecium Caudatam:
Paramecium Caudatum se reproduce asexualmente por fisión binaria transversal y también experimenta varios tipos de reorganización nuclear, como conjugación, endomixis, autogamia, citogamia y hemixis, etc.
(i) Fisión binaria transversal:
La fisión binaria transversal es el tipo más común de reproducción asexual en Paramecium. Es un proceso asexual claramente único en el que un espécimen completamente desarrollado se divide en dos individuos hijos sin dejar un cadáver parental.
El plano de división pasa por el centro de la célula y en un plano perpendicular al eje largo del cuerpo. La división del cuerpo celular en su conjunto va siempre precedida de la división de los núcleos; de hecho, parece que la reproducción se inicia con la actividad y división nuclear.

Paramecium Caudatum se reproduce por fisión binaria transversal en condiciones favorables. En la fisión binaria, el micronúcleo se divide por mitosis en dos micronúcleos hijos, que se desplazan a extremos opuestos de la célula. El macronúcleo se alarga y se divide transversalmente por amitosis.
Se brota otra citofaringe y aparecen dos nuevas vacuolas contráctiles, una cerca del extremo anterior y otra cerca del extremo posterior. Mientras tanto, aparece un surco de constricción cerca de la mitad del cuerpo y se profundiza hasta que el citoplasma está completamente dividido.
Los dos paramecios «hijos» resultantes son de igual tamaño y cada uno contiene un conjunto de orgánulos celulares. De los dos paramecios hijos producidos, el anterior se llama proter y el posterior se llama opisthe. Crecen hasta alcanzar su tamaño completo antes de que se produzca otra división.
El proceso de fisión binaria requiere unas dos horas para completarse y puede ocurrir de una a cuatro veces por día, produciendo de 2 a 16 individuos. Se producen unas 600 generaciones en un año.
La tasa de multiplicación depende de las condiciones externas de alimentación, temperatura, edad del cultivo y densidad de población; también de los factores internos de herencia y fisiología. Naturalmente, si todos los descendientes de un individuo sobrevivieran y se reprodujeran, el número de paramecios producidos pronto sería igual al volumen de la tierra.
El término clon se utiliza para referirse a todos los individuos que se han producido a partir de un individuo por fisión. Todos los miembros de un clon son hereditarios por igual.
(ii) Conjugación:
Ordinariamente el Paramecium Caudatum se multiplica por fisión binaria durante largos períodos de tiempo, pero a intervalos esto puede ser interrumpido por la unión de dos animales a lo largo de sus superficies orales para el proceso sexual de conjugación.
La conjugación se define como la unión temporal de dos individuos que intercambian mutuamente material micro nuclear. Es un tipo único de proceso sexual en el que dos organismos se separan poco después del intercambio de material nuclear.
Sonneborn (1947), basándose en el comportamiento de apareamiento de Paramecium Caudatum, ha informado de que cada especie de Paramecium existe en un número de variedades o singenes. Además, dentro de cada singén hay un número de tipos de apareamiento que suelen ser dos.
Los tipos de apareamiento siguen siendo morfológicamente idénticos pero presentan diferencias fisiológicas. En P. aurelia, hay 14 singenes y 28 tipos de apareamiento, mientras que en P. caudatum, hay 16 singenes y 32 tipos de apareamiento. Se ha observado que, por lo general, los paramecios no se conjugan con los miembros de su propio tipo de apareamiento ni con las demás variedades, sino sólo con el segundo tipo de apareamiento de su propia variedad.

Factores que inducen la conjugación:
Los factores que inducen la conjugación varían de una especie a otra, pero a continuación se indican algunos de ellos:
1. La conjugación se produce generalmente en condiciones de vida poco favorables; se dice que la inanición o la escasez de alimentos y una dieta bacteriana particular o ciertas sustancias químicas inducen el proceso de conjugación en ciertas especies de Paramecium.
2. La conjugación se produce después de unas 300 generaciones asexuales de fisión binaria, o se alterna con la fisión binaria a largos intervalos para rejuvenecer el clon moribundo, es decir, se produce en los individuos que deben haber pasado por un número deseable de generaciones asexuales, lo que se dice que es el período de inmadurez, y luego se vuelven sexualmente maduros para conjugarse.
3. La conjugación se produce cuando hay un cambio en la condición fisiológica de los paramecios, entonces se produce entre aquellos individuos que son algo más pequeños en tamaño (210 micras de largo) y se encuentran en una etapa que puede considerarse como un período de vejez insalubre; los paramecios de esta condición morirán si no se les permite conjugar.
4. Se dice que la oscuridad repentina en condiciones de luz y las bajas temperaturas inducen el proceso de conjugación en algunas especies.
5. La conjugación no tiene lugar durante la noche o la oscuridad; comienza a primera hora de la mañana y continúa hasta la tarde.
6. Se dice que una sustancia proteinácea en los cilios de los individuos del tipo de apareamiento induce la conjugación.
Proceso de Conjugación:
El proceso de conjugación difiere en las diferentes especies de Paramecium, pero el relato anotado se refiere al proceso de conjugación de P. caudatum (Fig. 20.21).
En la conjugación dos Paramecium caudatum (denominados preconjugantes) de los tipos de apareamiento opuestos de la misma variedad se juntan con sus superficies ventrales y se unen por sus surcos orales; sus cilios producen una sustancia en la superficie del cuerpo que provoca la adhesión de los dos paramecios conjugantes.
Dejan de alimentarse y su aparato de surcos orales desaparece. La película y el ectoplasma, en el punto de contacto, de ambos se rompen, y se forma un puente protoplásmico entre los dos animales. Ahora, estos individuos se llaman conjugantes.
En esta condición, la pareja conjugante nada activamente y simultáneamente se producen una serie de cambios nucleares en cada conjugante como se describe a continuación:
El macronúcleo comienza a desintegrarse, se afloja en su textura y forma una compleja madeja retorcida, durante la última mitad del período de conjugación desaparecerá finalmente siendo absorbido por el citoplasma. El micronúcleo de cada conjugante se divide dos veces, siendo una de ellas una división de reducción.
Así, en cada conjugante se producen cuatro micronúcleos hijos haploides. Tres de estos cuatro micronúcleos degeneran en cada uno, de modo que sólo queda uno.
El micronúcleo restante de cada conjugante se divide mitóticamente en dos pronúcleos o núcleos gaméticos desiguales formando un pronúcleo femenino estacionario más grande y un pronúcleo masculino migratorio más pequeño.
El pronúcleo migratorio de un conjugante cruza el puente protoplásmico y se fusiona con el pronúcleo estacionario del otro conjugante para formar un sincarion o núcleo de conjugación en el que se restablece el número diploide de cromosomas y se ha producido un intercambio de material hereditario.
El proceso se ha comparado con la fecundación en animales superiores, pero no se trata de una fecundación porque no hay gametos implicados. Los conjugantes ahora (después de unas 12-48 horas) se separan y se llaman ex-conjugantes. El sincarion de cada exconjugante se divide tres veces para formar ocho micronúcleos en cada exconjugante.
Cuatro de los ocho micronúcleos se agrandan y se convierten en macronúcleos, y tres de los otros cuatro micronúcleos desaparecen.
El micronúcleo restante se divide y al mismo tiempo el ex-conjugante se divide por fisión binaria en dos células, cada una de las cuales tiene dos macronúcleos y un micronúcleo. Las células y sus micronúcleos se dividen una segunda vez para formar cuatro paramecios a partir de cada exconjugante, de modo que cada uno tiene un macronúcleo y un micronúcleo.
El nuevo macronúcleo, como también el micronúcleo, han sido hechos de material nuevo. Estos nuevos núcleos contienen probablemente un potencial nuevo y diferente que se refleja en los individuos sanos.
Significado de la Conjugación:
Un clon se extinguirá si no se produce la reordenación nuclear, pero el clon puede rejuvenecerse para recuperar su antiguo vigor mediante la reordenación nuclear, esta reordenación nuclear se produce mediante la conjugación, por lo tanto, la conjugación es esencial para la continuidad de la fisión binaria.
La importancia de la conjugación se ha resumido a continuación:
1. La conjugación sirve como proceso de rejuvenecimiento y reorganización mediante el cual se restablece la vitalidad de la raza. Si la conjugación no se produce durante largos períodos, los paramecios se debilitan y mueren. (La afirmación de Woodruffs de mantener los paramecios sanos durante 22.000 generaciones sin conjugación es refutada por Sonneborn, porque demostró que todos los paramecios de Woodruffs pertenecían al mismo tipo de apareamiento).
2. No hay distinción de sexo en los conjugados aunque sólo los paramecios de dos tipos de apareamiento diferentes de la misma variedad se conjugarán.
3. No hay distinción de sexo, pero el pronúcleo migratorio activo se considera masculino y el estacionario femenino.
4. La conjugación es sólo una unión temporal, no hay fusión de citoplasma y no se produce ningún cigoto, pero el núcleo de cada ex-conjugante contiene material hereditario de dos individuos conjugados.
5. La conjugación conlleva la sustitución del macronúcleo por material procedente del sincarion, este es un acontecimiento de importancia fundamental. En la fisión binaria los cromosomas del macronúcleo se distribuyen al azar a las células hijas, la fisión binaria continuada ha hecho que el clon sea débil con algunas anormalidades estructurales.
La conjugación hace que se forme el número correcto de cromosomas en el macronúcleo, por lo que la raza se renueva en vigor. La función del micronúcleo es restablecer un complejo cromosómico y génico equilibrado.
Comportamiento aberrante en la reproducción en Paramecium Caudatum:
Paramecium Caudatum muestra ciertas variaciones en su comportamiento nuclear durante la fisión y la conjugación, estas desviaciones son endomixis, autogamia, citogamia y hemixis. En los tres primeros procesos se efectúa la recombinación genética y se forma un nuevo macronúcleo a partir del micronúcleo.
(i) Endomixis:
Woodruff y Erdmann (1914) informaron por primera vez de un nuevo proceso de reorganización nuclear, la endomixis (Gr., endon = dentro; mixis = mezcla) en Paramecium aurelia, una especie bimicronucleada (Fig. 20.22). Este proceso se describió como algo que ocurre periódicamente y en el que se produce un nuevo aparato macronuclear sin formación de sincariones. La endomixis se produce dentro de un mismo individuo.
Según Woodruff y Erdmann, el macronúcleo degenera y los micronúcleos se dividen dos veces para formar ocho micronúcleos. Seis micronúcleos degeneran y quedan dos. Con sólo dos micronúcleos restantes, el animal se divide por fisión en dos células, cada una con un micronúcleo.
El micronúcleo de cada célula hija se divide dos veces para formar cuatro micronúcleos. Dos micronúcleos se amplían para formar macronúcleos. El animal y sus micronúcleos se dividen de forma que se forman dos individuos hijos, cada uno con un macronúcleo y dos micronúcleos.
La endomixis se produce en aquella variedad de P. aurelia que no se conjuga, por lo que el efecto de la endomixis puede ser el mismo que el de la conjugación, ya que ambos procesos provocan la sustitución del macronúcleo por material procedente del micronúcleo, y ambos procesos rejuvenecen la vitalidad de la raza.
Pero los dos procesos difieren porque no hay fusión de pronúcleos en la endomixis; la endomixis puede compararse con la partenogénesis.
Sin embargo, algunos trabajadores han afirmado con buenas razones que la endomixis no es un proceso válido y que se ha descrito debido a una observación defectuosa. Con toda probabilidad, la endomixis no tiene lugar y puede ser sólo un caso especializado de autogamia.

Más tarde, Erdmamm y Woodruff (1916) informaron de la endomixis en Paramecium caudatum. Diller, sin embargo, no cree en la validez de este proceso y considera que Erdmann y Woodruff simplemente han combinado las etapas de hemixis y autogamia en un solo esquema, la endomixis.
(ii) Autogamia:
Diller (1934, 1936) y Sonneborn (1950) describieron un proceso de autofecundación o autogamia que ocurre en un solo individuo en Paramecium aurelia (Fig. 20.23). Informó que en la autogamia tres divisiones micronucleares (pregámicas), que implican la maduración, producen los núcleos gaméticos (pronúcleos).
Durante la autogamia en P. aurelia, los dos micronúcleos se dividen dos veces (una meióticamente) para formar ocho micronúcleos, seis de los cuales degeneran. Mientras tanto, el macronúcleo crece en una masa en forma de madeja que se rompe en pedazos para ser absorbida posteriormente en el citoplasma. Dos de los ocho micronúcleos, como pronúcleos, entran en un cono protoplásmico abultado cerca de la boca de la célula.
Los dos pronúcleos se fusionan para formar el sincarion. El sincarion se divide dos veces para formar cuatro micronúcleos. Dos micronúcleos se convierten en macronúcleos. El paramecio y sus micronúcleos se dividen para formar dos individuos hijos, cada uno con un macronúcleo y dos micronúcleos. Este proceso se completa en unos dos días.
La autogamia provoca el rejuvenecimiento de la raza.
Se asemeja a la conjugación en cuanto que el nuevo macronúcleo se forma con material del micronúcleo, en el nuevo macronúcleo se restablece el número correcto de cromosomas; y también en el hecho de que se produce la fusión de dos pronúcleos. Pero la autogamia se diferencia de la conjugación porque en la autogamia participa un solo individuo y aporta los dos pronúcleos, es una especie de autofecundación.
(iii) Hemixis:
Diller (1936) informó de la hemixis en Paramecium aurelia (Fig. 20.24). La hemixis es principalmente un proceso de fragmentación y división macronuclear sin ninguna actividad micronuclear inusual. Diller clasificó la hemixis en cuatro tipos, a saber, A, B, C y D, como se muestra en la Fig. 20.24 en P. aurelia, pero también encontró todos los tipos en cultivos masivos de P. caudatum y P. multimicronucleatum.
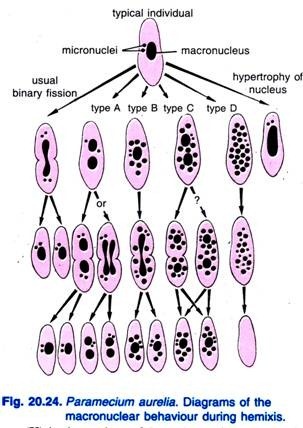
El tipo A es la forma más simple de hemixis caracterizada por una división del macronúcleo en dos o más partes. Esta división no está sincronizada con la división micronuclear.
El tipo B se caracteriza por la extrusión de una a 20 o más bolas de cromatina del macronúcleo al citoplasma.
El tipo C se caracteriza por la división simultánea del macronúcleo en dos o más porciones principales y la extrusión de bolas macronucleares al citoplasma.
El tipo D se considera que representa condiciones patológicas en las que el macronúcleo sufre una fragmentación completa en bolas de cromatina que finalmente desaparecen de la célula. Los micronúcleos suelen desaparecer antes de la disolución del macronúcleo.
(iv) Citogamia:
Wichterman (1939) ha informado de otro proceso sexual en Paramecium caudatum, que denominó citogamia. En la citogamia no hay intercambio nuclear. En este proceso, dos individuos se unen por sus superficies ventrales, pero la película de los dos individuos no se rompe.
El micronúcleo de cada individuo se divide tres veces para formar ocho micronúcleos, seis de los cuales se desintegran en cada individuo. Los dos micronúcleos restantes se fusionan para formar un sincarion en cada célula. Los animales se separan ahora.
La citogamia difiere de la autogamia en que hay dos animales en contacto entre sí, pero se asemeja a la autogamia y a la conjugación en la fusión de dos pronúcleos. La citogamia se diferencia de la conjugación en que no hay intercambio nuclear entre los dos animales que se unen.
Algunas partículas citoplasmáticas reportadas en Paramecium Caudatum:
(i) Partículas Kappa:
En 1938, T.M. Sonneborn informó que algunas razas (conocidas como asesinas o cepa asesina) de Paramecium producen una sustancia venenosa, llamada paramecina que es letal para otros individuos llamados sensibles. La paramecina es soluble en agua, difusible y depende para su producción de algunas partículas localizadas en el citoplasma del Paramecium (cepa asesina).
Estas partículas se denominan partículas kappa. Las partículas kappa tienen ADN y ARN. Un Paramecio asesino puede contener cientos de partículas kappa. El estudio detallado de estas partículas ha revelado que un gen dominante (K) en el núcleo del Paramecium es necesario para que las partículas kappa existan, se multipliquen y produzcan paramecina.
(ii) mµ, partículas:
R.W. Siegel (1952) informó de otro tipo de partículas asesinas en el citoplasma de algunos Paramecium. Un Paramecio con partículas mp se llama mate killer porque cuando se conjuga con un Paramecio sin partículas mµ llamado mate sensitive, entonces mata a este último. Las partículas mp también están compuestas por ADN, ARN, etc.
Estas partículas sólo existen en aquellos paramecios cuyo micronúcleo contiene al menos un gen dominante de cualquiera de los dos pares de genes cromosómicos no vinculados (M1 y M2).
(iii) Partículas Pi:
Estas partículas se supone que son la forma mutante de las partículas kappa pero no producen ningún tipo de sustancia venenosa.
(iv) Partículas Lambda:
Estas partículas se reportan en los paramecios asesinos y se dice que producen alguna sustancia responsable de causar la lisis o desintegración de los paramecios sensibles, es decir, que no la poseen.