Gametes og befrugtning oversigt
Kønslig reproduktion henviser til de seksuelle kønsceller (gameter), der produceres af forældrene, gennem kombinationen af biseksuelle kønsceller (såsom sædceller og ægceller), bliver til det befrugtede æg, og derefter udvikler det befrugtede æg sig til et nyt individ, kaldet kønslig reproduktion. Omfattende variation i genkombinationer ved seksuel reproduktion kan øge afkommets evne til at tilpasse sig til den naturlige udvælgelse. Tilfældigt kombinerede gener i afkommet fra seksuel reproduktion kan være til gavn for arten eller ej, men øger i det mindste chancerne for, at nogle få individer overlever i et uforudsigeligt og evigt foranderligt miljø, hvilket er til gavn for arten. Seksuel reproduktion kan også fremme spredningen af gunstige mutationer i populationen. Hvis to individer i en art har gunstige mutationer på forskellige steder, vil de to mutanter i den aseksuelle reproduktionspopulation konkurrere, indtil en udslettelse, det er umuligt at bevare begge gunstige mutationer på samme tid. I seksuelt reproducerende populationer kan disse to gunstige mutationer imidlertid gennem parring og rekombination samtidig komme ind i genomet hos det samme individ og samtidig spredes i populationen. Af ovennævnte grunde fremskynder seksuel reproduktion evolutionsprocessen. I den mere end 3 milliarder år lange biologiske udvikling på Jorden har de første to milliarder år af livet været i den aseksuelle reproduktionsfase, og udviklingen har været langsom, mens udviklingshastigheden er accelereret betydeligt siden den sidste 1 milliard år. Ud over ændringer i jordens miljø (såsom fremkomsten af den iltholdige atmosfære osv.) er forekomsten og udviklingen af seksuel reproduktion også en væsentlig årsag.
Gametes
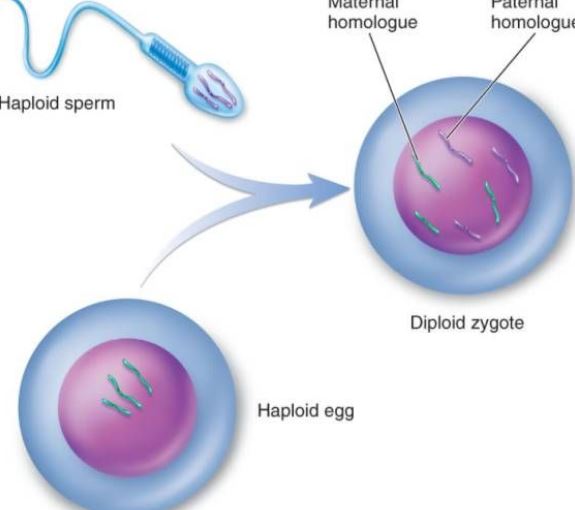
Gametes henviser til modne celler, der produceres af forplantningssystemet, når organismen er seksuelt reproducerende, benævnt kimceller. Gametes opdeles i hanlige kønsceller og hunlige kønsceller. De kvindelige kønsceller hos dyr og planter kaldes normalt ægceller, og de mandlige kønsceller kaldes sædceller. Sædcellerne er ret små, men de kan bevæge sig og trænge ind i ægcellen i skorpionform. Ægcellen er ret stor og ikke-svømmende. F.eks. er ægcellens volumen hos søpindsvin 10.000 gange større end sædcellens volumen. Selv om han- og hunkønslegemer er forskellige i volumen, er det kernedNA, de leverer til afkommet, lige stort, dvs. de leverer hver især et sæt genomer. På grund af ægcellernes store størrelse leveres dattercellernes cytoplasmiske struktur og cytoplasmiske DNA imidlertid grundlæggende af ægcellerne. Gameterne spiller en vigtig rolle i biologiske beregninger. Ved hjælp af genetiske kort kan genstrømmene og mønsteret af afkomsgenotyperne observeres tydeligt. Antallet af gameter i den første generation af hybrider og de relative egenskaber i hybrider er 2n-te potens (n er logaritmen af relative egenskaber). Når et levende legeme danner en gamet, adskilles de parrede genetiske faktorer fra hinanden og indgår i forskellige gameter. Gameterne indeholder kun ét af hvert par af genetiske faktorer. Når de befrugtes, er kombinationen af han- og hun-gameter tilfældig. På grund af slægten af gameter, der dannes ved meiose, mangfoldigheden i kromosomsammensætningen, forskellen i genetisk materiale i forskellige gameter og tilfældigheden i kombinationen af ægceller og sædceller i befrugtningsprocessen, må afkommet af samme forælder være forskelligartet.
Fertilisation
Fertilisation er den proces, hvorved æg og sædceller smelter sammen til en zygote. Den er et grundlæggende træk ved seksuel reproduktion og er allestedsnærværende i dyre- og planteverdenen, men den er ofte omtalt som den mest omtalte. Dyrs befrugtning på celleniveau omfatter befrugtningsprocessen tre hovedstadier: aktivering af æggene, regulering og amfoterisk pronuklear fusion. Aktivering kan betragtes som ontogeniens startpunkt, der hovedsageligt manifesteres ved ændringer i ægmembranens permeabilitet, ekklesi af kortikale granula, dannelsen af den befrugtede membran osv. Justering sker efter aktivering, som er det første skridt til at sikre den normale deling af befrugtede æg. Ændringen af den prokaryotiske kerne sikrer arven fra forældrene og genopretter den diploide kerne. Befrugtningen sætter ikke kun gang i replikationen af DNA’et, men aktiverer også den genetiske information såsom mRNA og rRNA i ægget for at syntetisere det protein, der er nødvendigt for embryonets udvikling. Efter 6 til 7 dage efter befrugtningen forsvinder blastocysten zona pellucida og begraves gradvist og dækkes af endometrium, hvilket kaldes implantation af det befrugtede æg. Sædcellerne hos et dyr har ikke tydelig kemotaxis som sædcellerne hos lavere planter som f.eks. bryofytter, men når snarere i nærheden af ægget ved aktiv bevægelse eller ved ciliær bevægelse af de genitale epithelceller. Kapacitering af sædceller: Det er kendt, at når mange pattedyrs sædceller passerer gennem de kvindelige kønsorganer eller gennem cumulus, fjernes det fremmede protein, der indkapsler sædcellerne, og de fysiske og biologiske egenskaber af sædcellernes plasmamembran ændres, hvilket gør det muligt for sædcellerne at få energi og deltage i befrugtningsprocessen. Når pattedyrs-sædcellen med kapacitet kommer i kontakt med ægmembranen eller zona pellucida på ægget, binder den specifikt til et glykoprotein på æggemembranen, hvilket stimulerer sædcellen til at producere energi. Akrosomreaktionen hjælper sædcellerne til at krydse æggemembranen yderligere. På søpindsvinets æg er sædcellens akrosomreaktion et bestemt polysaccharidstof i peri-membranfilmen. De fleste æg har æggemembraner i periferien, og tykkelsen af de forskellige æggemembraner varierer. Hovedbestanddelen er mucin eller mucopolysaccharid; kun få er nøgne æg, som f.eks. æggene fra en coelenterat. Acrosomreaktion: Når sædcellen passerer gennem æggemembranen, sker der en bindingsproces. Førstnævnte er løst knyttet, fri for ydre temperaturinterferens og har ingen specificitet. I løbet af adhæsionsperioden omdannes protoplastproteinet på den akrosomale membran til akrosomprotein, og akrosomproteinet fremskynder sædcellerne til at krydse æggemembranen; Det er en stærk kombination, kan forstyrres af lav temperatur og har en specificitet. Der er blevet isoleret et protein, der binder specifikt til ægglykoproteinet på plasma-membranen hos søpindsvinssædcellerne, kaldet et bindingsprotein, med en molekylvægt på ca. 30.000. Det er en stærk styrke, der kan forstyrres af den stærke temperatur, og det har en specificitet. De sekundære ægceller i midten af den anden modningsdeling under ægløsningen trænger sammen med den omgivende zona pellucida og strålekronen hurtigt ind i æggelederampullen gennem bughulen på grund af svingningen af cilia i æggelederepitelcellerne og sammentrækningen af muskulaturlaget. Hvis ægcellen ikke kan møde sædcellen, begynder den normalt at degenerere inden for 12-24 timer. Når først sædcellen er i kontakt med ægget, gennemgår selve ægget en række aktiveringsændringer. På pattedyrsæg er den karakteriseret ved kortikalreaktionen, ovariehinde-reaktionen og zona pellucida-reaktionen, som kan blokere flerfoldsbefrugtningen og stimulere den videre udvikling af æggene. Den kortikale reaktion forekommer på tidspunktet for sædcellens fusion. Fra fusionstidspunktet brister de kortikale partikler, og indholdet udledes, hvorved hele æggets cortex spredes. Æggemembranreaktionen er en rekombinationsproces af ægget og den kortikale kornhinde. Zona pellucida-reaktionen er en proces, hvor kortikaludstrømningen og zona pellucida fra en befrugtet membran, æggemembranen adskilles fra plasmamembranen, sædreceptoren i zona pellucida forsvinder, og zona pellucida hærder. Kun sædceller, der har en akrosomreaktion, kan fusionere med ægget. Under påvirkning af acrosin passerer sædcellen gennem den radiale krone og interagerer med sædreceptor-glykoproteinmolekylet ZP3 på zona pellucida, hvilket gør det muligt for sædcellen at frigive det akrosomale enzym og trænge ind i perivitellinen gennem zona pellucida. I begyndelsen af befrugtningen er omslaget af den ækvatoriale del af det menneskelige sædhoved i kontakt med ægcellemembranen, og derefter trænger sædcellens kerne og cytoplasma ind i ægget. Efter at sædcellen er kommet ind i ægget, frigiver de kortikale partikler i æggets lavvandede cytoplasma straks sit indhold til membranens periferi. Samtidig øges den negative overfladeladning på celleoverfladen i takt med, at de kortikale granulers membran smelter sammen med ægcellerne, hvorved sædcellernes plasmamembran undertrykkes i fusionen med æggemembranen, hvilket kaldes en kortikal reaktion. Zona pellucida-strukturen ændrer sig, hvilket kaldes zona pellucida-reaktionen. På dette tidspunkt reduceres zona pellucida’s evne til at binde sædceller, hvilket forhindrer forekomsten af polyspermy og sikrer de biologiske karakteristika ved menneskelig monospermy. Ud over at ødelægge eller inaktivere den receptor, der er bundet til sædcellerne i zona pellucida, kan indholdet af de kortikale granula også øge tværbindingen mellem peptidkæderne i zona pellucida, svække følsomheden over for acrosomprotease og forhindre sædgennemtrængning. Selv om der er flere sædceller, der passerer gennem zona pellucida, er der kun én sædcelle, der trænger ind i ægcellen for at befrugte den. I en unormal situation er der to sædceller involveret i befrugtningen, nemlig dobbelt sædbefrugtning. To sædceller trænger samtidig ind i ægget og danner embryoner af triploide celler, som alle aborteres eller dør kort efter fødslen. Efter at sædcellerne er kommet ind i ægget, gennemfører ægget hurtigt den anden modne deling. På dette tidspunkt kaldes sæd- og ægkernerne for henholdsvis den mandlige pronucleus og den kvindelige pronucleus. De to pronuclei nærmer sig gradvist hinanden, kernemembranen forsvinder, og kromosomerne smelter sammen og danner et diploidt befrugtet æg. Når sædcellerne er smeltet sammen, kan man se, at mikrovilli på æggets overflade omgiver sædcellen, som kan være orienteret; derefter smelter æggemembranen sammen med plasmamembranen i sædcellens acrosom i det bageste område.
Reference
- Yanagimachi R. Germ cells and fertilization: Why I studied these topics and what I learned along the path of my study. Andrologi. 2014, 2(6):787-93.
- Henshaw J M, Marshall D J, Jennions M D, et al. Local gamete competition explains sex allocation and fertilization strategies in the sea. American Naturalist. 2014, 184(2):32.
- Mori T, Igawa T. Gamete attachment process revealed in blooming plant fertilization. Plant Signaling & Behavior. 2014, 9(12):977715.
- Hédouin L, Pilon R, Puisay A. Hyposalinitetsstress kompromitterer befrugtningen af gameter mere end korallelarvernes overlevelse. Marine Environmental Research. 2015, 104:1-9.
Luo J, Mcginnis L K, Carlton C, et al. PTK2b-funktion under befrugtning af musens oocyt. Biochem Biophys Res Commun. 2014, 450(3):1212-1217.