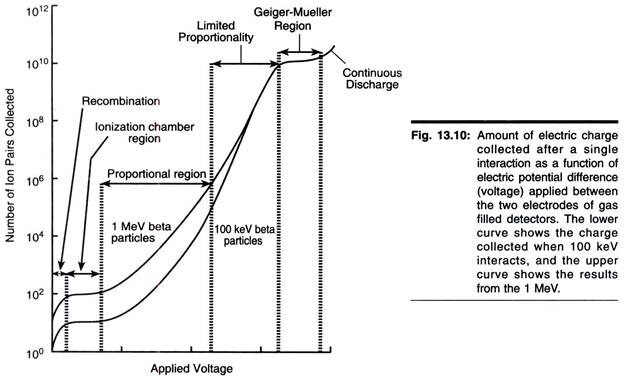
ADVERTISEMENTS:
I denne artikel vil vi diskutere Paramecium Caudatum:- 1. Levested, levested og kultur af Paramecium Caudatum 2. Struktur af Paramecium Caudatum 3. 3. Bevægelighed 4. Ernæring 5. Respiration og udskillelse 6. Osmoregulering 7. Adfærd 8. Reproduktion 9. Afvigende adfærd i forbindelse med reproduktion 10. Nogle cytoplasmatiske partikler.
Indhold:
- Habit, Habitat og kultur af Paramecium Caudatum
- Struktur af Paramecium Caudatum
- Lokomotion af Paramecium Caudatum
- Næring hos Paramecium Caudatum
- Respiration og udskillelse hos Paramecium Caudatum
- Osmoregulering af Paramecium Caudatum
- Adfærd hos Paramecium Caudatum
- Fremstilling hos Paramecium Caudatum
- Afvigende adfærd ved forplantning hos Paramecium Caudatum
- Nogle cytoplasmatiske partikler rapporteret hos Paramecium Caudatum
- Habit, Levested og dyrkning af Paramecium Caudatum:
- Kultur af Paramecium:
- Struktur af Paramecium caudatum:
- (i) Størrelse og form af Paramecium caudatum:
- (ii) Pellicle:
- (iii) Cilia:
- (iv) Infra-ciliært system:
- (v) Mundrille og cytopyge:
- (vi) Cytoplasma:
- (vii) Ektoplasma:
- (viii) Trichocyster:
- (ix) Neuromotorium og tilknyttede fibriller:
- (x) Endoplasma:
- (xi) Kontraktile vakuoler:
- (xii) Fødevakuoler:
- Lokomotion hos Paramecium Caudatum:
- (i) Metaboly eller kropsforvridninger:
- (ii) Ciliær lokomotion:
- Næring hos Paramecium Caudatum:
- Næringsmekanisme:
- Forrådnelse og ætsning hos Paramecium caudatum:
- Respiration og udskillelse af Paramecium Caudatum:
- Adfærd hos Paramecium Caudatum:
- (i) Reaktioner på kontakt (Thigmotaxis):
- (ii) Reaktioner på kemikalier (kemotaksi):
- (iii) Reaktioner på temperatur (Thermotaxis):
- (iv) Reaktioner på lys (Phototaxis):
- (v) Reaktioner på elektrisk strøm (Galvanotaxis):
- (vi) Reaktioner på vandstrøm (Rheotaxis):
- (vii) Reaktioner på tyngdekraften (Geotaxis):
- Fremstilling hos Paramecium Caudatam:
- (i) Tværgående binær fission:
- (ii) Konjugation:
- Afvigende adfærd i reproduktionen hos Paramecium Caudatum:
- (i) Endomixis:
- (ii) Autogami:
- (iii) Hemixis:
- (iv) Cytogami:
- Nogle cytoplasmatiske partikler rapporteret i Paramecium Caudatum:
- (i) Kappa-partikler:
- (ii) mµ, Partikler:
- (iii) Pi-partikler:
- (iv) Lambdapartikler:
Habit, Levested og dyrkning af Paramecium Caudatum:
Paramecium caudatum (Gr., paramekes = aflang; L., caudata = hale) er almindeligt forekommende i ferskvandsdamme, bassiner, grøfter, vandløb, søer, reservoirer og floder. Den findes især i rigelige mængder i stillestående damme, der er rige på rådnende materiale, i organiske opblandinger og i spildevand. Paramecium caudatum er en fritlevende organisme, og denne art er udbredt over hele verden.
Kultur af Paramecium:
ADVERTISEMENTS:
Tag nedsænket ukrudt fra en dam og læg det i en krukke med destilleret vand, dæk krukken til og lad den rådne; der vil komme sværme af Paramecia i løbet af få dage. Kog nu hø i vand, dekanter infusionen og tilsæt et par hvedekorn, og lad den stå indtil den er grumset af bakterier.
Overfør Paramecia fra det første glas til denne væske, hvor de vil formere sig hurtigt. Høinfusioner alene vil producere Paramecia, der viser tilstedeværelse af cyster, og der er rapporteret om cyster, der ligner sandkorn, men der er intet bevis for, at Paramecia danner cyster, da de aldrig er blevet bekræftet.
Struktur af Paramecium caudatum:
(i) Størrelse og form af Paramecium caudatum:
Paramecium caudatum (Fig. 20.1) er en mikroskopisk organisme og er synlig for det blotte øje som et meget lille aflangt legeme.
ADVERTISEMENTS:
Den fremstår lysegrå eller hvid og måler almindeligvis mellem 170 og 290 mikrometer i længden og kan nå en længde på op til 300-350 mikrometer. P. caudatum ligner sålen på en tøffel eller en sko, og derfor er dyret almindeligvis kendt som tøfedyr. Den er fire gange så lang som bred og noget cylinderformet med tydeligt forskellige ender.
Den forreste del, der bevæger sig fremad, er slank med en stump eller afrundet ende, mens den bageste ende er noget spids eller kegleformet. Den bredeste del af organismen er lige under midten. Dyrets krop er asymmetrisk i form med en veldefineret oral eller ventral overflade og en aboral eller dorsal overflade.
(ii) Pellicle:
Kroppen er dækket af en tynd, dobbeltlaget, elastisk og fast pellicle af gelatine. Pelliclen holder dyrets form, men er elastisk nok til at tillade sammentrækninger. Pelliclen har dobbelt membran, den ydre membran er sammenhængende med cilierne og den indre membran med ektoplasmaet. I højere forstørrelse af mikroskopet viser pelliclen rektangulære eller sekskantede fordybninger på sin overflade.

Dette arrangement er bevaret på rygfladen af Paramecium, men på ventralfladen konvergerer kammene foran og bagved mod en præoral og postoral åbning. Hver sekskantet fordybning er perforeret af en central åbning, hvorigennem et enkelt cilium træder ud. De forreste og bageste rande af de sekskantede fordybninger bærer åbningerne til trikocyster.
Ehret og Powers’ (1957) elektronmikroskopiske undersøgelse af pellicle (fig. 20.2) har afsløret, at de sekskantede fordybninger svarer til regelmæssige serier af hulrum, alveolerne. Alle alveolerne danner tilsammen et sammenhængende alveolærlag, som er afgrænset af en ydre alveolær og en indre alveolærmembran.
Det ydre lag ligger i tæt kontakt under den ydre cellemembran. Derfor omfatter pellicle den ydre cellemembran, den ydre alveolarmembran og den indre alveolarmembran.
(iii) Cilia:
Hele kroppen er dækket af talrige, små, hårlignende fremspring kaldet cilia. Cilia forekommer i langsgående rækker over hele kroppen, denne tilstand er kendt som holotrichous, hvor kroppens cilia er lige store. Cilia har samme struktur som flageller, de har en ydre protoplasmatisk kappe eller plasmamembran med ni dobbelte langsgående fibriller i en perifer ring. Hos nogle cilia er de ni ydre fibriller ikke parvis.
ADVERTISERINGER:
Der er to centrale langsgående fibriller, som er tyndere end de ydre fibriller. Hvert cilium udspringer af en basal granulat eller kinetosom. De ni par perifere fibriller smelter sammen og danner kinetosomets væg, således er kinetosomet et rør, som enten er åbent eller lukket i den nederste ende, de to centrale fibriller stopper i niveau med pelliclen hos de fleste ciliater.
Udgående fra kinetosomet er der en tynd rhizoplast, som ikke slutter sig til kernen. Mange Metazoa har også cilia, deres struktur er den samme, bortset fra at basalgranulatet er anderledes, og det har fine filamenter eller rodfibre, der strækker sig ned i cytoplasmaet. Men cilier adskiller sig fra flageller ved generelt at være mere talrige og kortere i størrelse.
Ciliaturen kan med fordel opdeles i krops- eller somatiske cilier, som findes på kropsoverfladen, og i orale cilier, som er knyttet til mundregionen. Kropscilierne er lige store, men de er længere i den bageste ende, deraf navnet caudatum. Cilia er organeller til at bevæge sig og indsamle føde, og de fungerer også som sensoriske receptorer og registrerer stimuli fra det ydre miljø.

Ultrastruktur af cilier:
Cilier og flageller har en fibrillær sammensætning. Ved basen har ciliet en diameter på ca. 0,2 mikrometer eller 2.000 A0, som kan være op til 10 mikrometer over celleoverfladen. Cilierne er afgrænset af en enhedsmembran med en tykkelse på 90 A0, som ligner og er kontinuerlig med plasmamembranen. Ciliums afgrænsede rum indeholder en vandig substans, der kaldes matrix.
I matrixen forbliver der indlejret elleve langsgående fibriller eller mikrotubuli. Ud af de elleve fibriller er to placeret i midten, mens de resterende ni fibriller forbliver anbragt perifert omkring de centrale fibriller. Hver af de ni ydre fibriller har en diameter på 360A0 og består af to subfibriller med en diameter på 180 til 250A0.
Disse underfibriller betegnes som subfibriller A og subfibriller B.
ADVERTISEMENTER:
Subfibrillen A er lidt større end subfibrillen B. Subfibrillen A giver to tykke fremspring eller arme ud fra sin ene side. Armene fra subfibrillen A på alle de ydre fibriller forbliver rettet i urets retning. Desuden ligger subfibrillen A tættere på ciliumets centrum end subfibrillen B. Begge subfibriller har en fælles væg med en tykkelse på 50A0.
De to centrale fibriller har ikke parvise subfibriller som de perifere ni fibriller, men indeholder hver kun en enkelt tubulus. Hver central fibrille har en diameter på ca. 250A0 og består af en 60A0 tyk væg.
Både de centrale fibriller forbliver adskilt af et rum på 350A0 og forbliver indhyllet i en fælles kappe. Gibbnos (1967) har rapporteret, at kappen af de centrale fibriller afgiver ni radialt orienterede led eller eger til hver underfibrille A.

Den højopløselige elektronmikroskopi har afsløret, at hver af de perifere og centrale fibriller af cilia og flageller er sammensat af ti til tolv filamenter af 40A0 tykkelse. Hvert filament er perleformet. Hver perle forbliver anbragt i gitter på 40 gange 50A0 i planet af tubulens væg. Disse perler betragtes som den grundlæggende underenhed i tubulusstrukturen.
(iv) Infra-ciliært system:
Det infra-cillære system er placeret lige under de pelliculære alveoler. Det består af kinetosomet eller basallegemet og kinetodesma. Cilia udspringer af kinetosomer, og fra hvert kinetosom udspringer en delikat cytoplasmatisk fibrille kaldet kinetodesma, (Fig. 20.2).
Under pelliculaen lidt til højre, men forbundet med alle kinetosomer i én langsgående række, ligger et langsgående bundt af flere kinetodesmata, et kinetodesmata fra hvert kinetosom strækker sig et stykke fortil ind i sit eget bundt af kinetodesmata.
En longitudinal række af kinetosomer med deres kinetodesmata danner en longitudinal enhed kaldet en kinety. Alle kineties eller kinetia udgør det infraciliære system hos en ciliat. Kinetierne ligger i cortexen under pellicula, og deres antal er ret konstant for hver ciliat.
Infraciliærsystemet styrer og koordinerer ciliernes bevægelser, og det bevirker dannelsen af organeller ved celledeling, f.eks. danner nogle kinetia munden. Ved binær fission hos ciliater skæres kinetierne på tværs i to, der hver går til én dattercelle, dette kaldes perikinetal fission.
(v) Mundrille og cytopyge:
På den ventrolaterale side er der en stor skrå, lavvandet fordybning kaldet mundrille eller peristom, som giver dyret et asymmetrisk udseende. Den løber skråt bagud fra den ene side (normalt fra venstre mod højre, men i nogle tilfælde fra højre mod venstre) og slutter lidt bag mellemkroppen. Mundrillen munder ud i en kort kegleformet tragtformet fordybning kaldet vestibulum.
ADVERTISER:
Vestibulummet fører direkte ind i den faste, ovalformede åbning kaldet cytostom (mund). Direkte fra cytostomet mod kroppens midte strækker sig det brede cytopharynx sig direkte ud fra cytostomet mod kroppens midte. Cytopharynx drejer derefter skarpt mod bagsiden for at blive til det slanke, tilspidsede spidse oesophagus.
Så er spiserøret nogenlunde parallelt med Parameciums kropsoverflade, undtagen ved den bageste ende. Her drejer spiserøret igen ind mod dyrets centrum for at føre ind i den dannende fødevakuole.
Cytopygen (også kaldet celleanus eller analplet eller cytoprokt) ligger på den ventrale overflade af kroppen næsten lodret bag cytostomet eller munden. Ufordøjede fødepartikler udskilles gennem cytopygen. Cytopharynx’ ciliation er meget kompliceret.
Gelei (1934) rapporterede om tilstedeværelsen af fire rækker, og Lund (1941) observerede mindst fire rækker. En struktur kaldet penniculus findes på cytopharynx’ venstre væg og spiralerer ca. 90 grader, så dens bageste ende befinder sig på den orale (ventrale) overflade af spiserøret.
I henhold til Lund består penniculus af otte rækker af cilier, der er arrangeret i to tætliggende blokke af hver. Et lignende bånd bestående af fire rækker lange cilier, der er mindre kompakt end i penniculus, betegnes som quadrulus. Det snor sig ned langs den dorsale væg i bughulen og ender tæt på penniculus.
Penniculus og quadrulus er fejlagtigt blevet kaldt en bølgende membran af nogle arbejdere. Quadrulus og penniculus kontrollerer passagen af føde. Det vides ikke, hvordan cilia virker, sandsynligvis trækker deres fibriller sig sammen på en rytmisk måde, som forårsager bøjning. Gelei (1925) påpegede, at penniculus’ funktion er at tvinge fødeelementer ind i kroppen.

(vi) Cytoplasma:
Cytoplasmaet er differentieret i en smal, ydre eller kortikal zone kaldet ektoplasmaet og en større, indre eller medullær region kaldet endoplasmaet.
(vii) Ektoplasma:
Ektoplasmaet (ektosarc eller cortex) er en permanent del af kroppen, der er markant afgrænset fra endoplasmaet. Ektoplasmaet danner et fast, klart, tyndt og tæt ydre lag. Det indeholder trikocyster, cilier og fibrillære strukturer og er udvendigt afgrænset af en belægning kaldet pellicle.
(viii) Trichocyster:
Indlejret i ektoplasmaet vinkelret på overfladen findes små spindelformede poser, der kaldes trikocyster. En lille plet på hver forreste og bagerste kant af sekskanten markerer placeringen af en trikocyst. De er fyldt med en brydningsfri, tæt væske med en svulmende substans, og i den ydre ende er der et kegleformet hoved eller en spids.
Trichocysterne ligger vinkelret i ektoplasmaet, de åbner sig ved små porer på højderyggene af de sekskantede områder af pelliclen.
De opstår fra kinetosomer af cilier, vandrer derefter og placerer sig i lige stor afstand i endoplasmaet. Når dyret irriteres, udstøder trichocysterne sig som lange klæbrige tråde. En udløst trichocyst har en uigennemsigtig spids, der ligner et omvendt søm, og et langt stribet skaft, men skaftet ses ikke i uudløst tilstand og er sandsynligvis dannet under udløsningen.
Trichocysternes funktion er usikker, men de udstøder sig som en reaktion på lokale kontakter og skader, de kan fungere som forsvarsorganeller.
Men dette er usikkert, fordi trichocysterne er ineffektive mod Didinium, det vigtigste rovdyr på Paramecium, de kan være til at fiksere dyret til et sted under fodring. Hos nogle ciliater fungerer trichocysterne som angrebsorganeller. Efter at trichocysterne er udledt, regenereres de fra kinetosomer.
Paramecium

(ix) Neuromotorium og tilknyttede fibriller:
I henhold til Lund (1933) findes der på venstre dorsalvæg af cytopharynx omtrent i højde med cytostomets bagrand en meget lille, tosidet masse, neuromotorium, på venstre side af cytopharynx. Fra neuromotorium udstråler fibriller ind i endoplasmaet.
Af disse går fire eller flere normalt næsten til den dorsale kropsvæg, men resten er kortere og ikke bestemte i deres placering. Alle betegnes som endoplasmatiske fibriller. Deres funktion er ukendt, men de koordinerer muligvis de orale cilieres fødebevægelser. Fibrillerne kan også give mekanisk støtte, elasticitet, kontraktilitet, ledningsevne og metabolisk indflydelse.

(x) Endoplasma:
Endoplasmaet eller medulla er den mere flydende og voluminøse del af cytoplasmaet, som indeholder mange cytoplasmiske granula samt andre inklusioner og strukturer af specialiseret karakter. De cytoplasmatiske inklusioner er mitokondrier, Golgi-apparater, vacuole, krystaller, granula og chromidier osv. Andre strukturer, dvs. kerner, kontraktile vacuoler og fødevakuoler, findes også i endoplasmaet.
Nuclei:
I endoplasmaet nær cytostomet findes to kerner, dvs. Paramecium er heterokaryote, en stor ellipsoid og granulær makronukleus og en anden lille kompakt mikronukleus.
Makronukleus er et iøjnefaldende, ellipsoidisk eller nyreformet legeme. Det er af en kompakt type, der indeholder fine tråde og tæt pakkede diskrete kromatinkorn af varierende størrelse og er indlejret i en achromatisk matrix. Den har mange nukleoler og meget mere kromatinmateriale (DNA).
Det er en somatisk eller vegetativ kerne. Den deler sig amitotisk og styrer dyrets vegetative funktioner (metaboliske aktiviteter). Den gennemgår dog mitose.
Micronukleus er lille, kompakt og kugleformet. Den findes generelt tæt på makronukleus ofte i en konkavitet. Fine kromatinkorn og -tråde er jævnt fordelt i hele strukturen. Mikronukleus deler sig mitotisk og kontrollerer reproduktionen.
Tallet af mikronukleus varierer med arten; det er ét hos P. caudatum, to hos P. aurelia og mange hos P. multimicronucleatum. Mikronucleus indeholder en tydelig nucleolus hos P. aurelia, men den findes ikke hos P. caudatum. Moses (1949; 1950) har rapporteret, at makronukleus og mikronukleus er identiske med hensyn til kemisk sammensætning.
(xi) Kontraktile vakuoler:
Der er to store, væskefyldte kontraktile vacuoler, som hver er placeret nær den ene ende af kroppen tæt på rygfladen. Deres placering er fast (i modsætning til Amoeba), de ligger mellem ektoplasmaet og endoplasmaet, men de er midlertidige organeller, der forsvinder med jævne mellemrum. Hos nogle arter ser de ud til at have en foringsmembran, i hvilket tilfælde de ikke forsvinder helt under systolen.
I forbindelse med hver kontraktile vacuole findes fem til tolv rørformede udstrålende kanaler, der hver består af en terminal del, en lang ampulla, der falder sammen, når den er tom, og en kort injektorkanal, der åbner sig i vacuolen.

Kanalerne kommunikerer med en stor del af kroppen, hvorfra de optager væsker og hælder dem i vacuolen, der således gendannes og vokser i størrelse, når den kontraktile vacuole når sin maksimale størrelse, trækker den sig pludselig sammen (systole) og udleder sit indhold gennem en permanent pore i pelliclen, derefter danner kanalerne igen de kontraktile vacuoler, kanalerne forsvinder ikke helt, da de er permanente strukturer.
De to kontraktile vacuoler udleder uregelmæssigt, den bageste kontraherer hurtigere, fordi den er tæt på cytopharynx, og der kommer mere vand ind i den. Hovedfunktionen af kanalerne og de kontraktile vacuoler er hydrostatisk, de fjerner overskydende vand fra protoplasmaet, vandet absorberes delvist og indtages delvist under fødslen.
Det kvælstofholdige affaldsstof har ammoniakforbindelser og nogle urater, som udstødes fra de kontraktile vacuoler sammen med CO2, men der er ingen tegn på, at udskillelsesstoffet udskilles af protoplasmaet i kanalerne eller de kontraktile vacuoler.
Der er en fast permanent cytopyge eller anus under peristomet til den ene side af cytopharynx, hvorigennem ufordøjede madrester og urater udstødes.
Den elektronmikroskopiske undersøgelse af kontraktile vacuoler har afsløret, at hvert kontraktile apparat består af nogle af tubuli af endoplasmatisk retikulum, nephridiske tubuli, feederkanaler, accessoriske vacuoler (radiale kanaler) og kontraktile hovedvakuoler. De accessoriske vacuoler formodes at være ampuller af feederkanaler (Fig. 20.11).
(xii) Fødevakuoler:
Disse er nogenlunde kugleformede, ikke-kontraktile legemer, der varierer i størrelse og antal, og som ligger i endoplasmaet. De indeholder optagne fødepartikler, hovedsagelig bakterier og en lille mængde væske, der er afgrænset af en tynd, tydelig membran. Volkonsky (1934) foreslog navnet gastrioler for disse vacuoler. I tilknytning til fødevakuolerne findes fordøjelsesgranulerne.
Lokomotion hos Paramecium Caudatum:
Paramecium Caudatum udfører lokomotion ved hjælp af to metoder, nemlig metabolisme eller kropsforvridninger og ved hjælp af cilier.
(i) Metaboly eller kropsforvridninger:
Kroppen hos Paramecium caudatum har elasticitet, den kan klemme sig gennem en passage, der er smallere end dens krop, hvorefter kroppen igen antager sin normale form. Denne midlertidige ændring af kroppens form er metabolisme, den fremkaldes i Paramecium af protoplasmaet.
(ii) Ciliær lokomotion:
Lokomotion fremkaldt af cilia er den vigtigste metode. Cilia kan slå fremad eller bagud, hvilket gør det muligt for dyret at svømme fremad eller bagud.
Normalt svømmer dyret fremad, idet cilierne slår bagud, men skråt, cilierne stivner og bøjer hurtigt bagud, så de næsten berører kropsoverfladen, dette kaldes det effektive slag; derefter bliver cilierne slappe og vender langsomt tilbage til den oprindelige lodrette position, dette kaldes genopretningsslag.
Cilia i samme tværgående række slår sammen, og dem i samme langsgående række slår efter hinanden fra den forreste til den bageste ende.

Denne koordinerede bevægelse af cilier kaldes metakron rytme, som skyldes det infraciliære system; dette bevirker, at dyret svømmer fremad. Men når kropscilia slår skråt bagud, så slår samtidig de længere cilia i mundrillen kraftigere, hvilket får den forreste ende til at svinge til venstre.
Kommunikationen mellem kroppens og mundrillenes cilia får dyret til at rotere om sin længdeakse. Denne rotation er altid til venstre (undtagen hos P. calkinsi, som roterer i en højrespiral).
Denne kombination af fremadgående bevægelse, svingning og rotation får dyret til at bevæge sig fremad i en spiralbane mod urets retning. Denne bane har en lige akse, og den samme kropsoverflade på dyret forbliver mod spiralbanens akse. Men når de svømmer baglæns, roterer alle arter til højre.
Ciliarslaget kan vendes om, således at cilierne bevæger sig skråt fremad, hvorved dyret svømmer baglæns. Ved hjælp af ciliernes bevægelse bevæger Paramecium sig med en hastighed på 1500 mikrometer eller endnu mere pr. sekund.
Jennings hævdede, at Parameciums spiralformede bevægelse skyldes, at mens cilierne hovedsageligt slår bagud, gør de det skråt til højre og får derved dyret til at rulle over til venstre.
Også denne slingring af kroppen mod den aborale overflade skyldes i høj grad den større kraft af det effektive slag af de orale cilier, som slår mere direkte bagud. Resultatet – Parameciums rotation om sin lange akse – gør det herved muligt for Paramecium at følge et mere eller mindre lige forløb ved dannelsen af store spiraler.
Næring hos Paramecium Caudatum:
I Paramecium Caudatum er ernæringen holozoisk. Føden består hovedsageligt af bakterier og små protozoer. Paramecium venter ikke på føden, men jager aktivt efter den.
Det hævdes, at Paramecium Caudatum udviser et valg i udvælgelsen af sin føde, men der synes ikke at være noget grundlag for dette, selvom den kun opsluger visse typer bakterier; de tilgængelige data tyder på, at 2 til 5 millioner individer af Bacillus coli fortæres af et enkelt Paramecium på 24 timer. Den lever også af encellede planter som alger, kiselalger osv. og små dele af dyr og grøntsager.
Næringsmekanisme:
Når Paramecium Caudatum kommer ind i et område med rigeligt med føde, kommer den til hvile. Den spiser kun i hvile eller når den svømmer meget langsomt, den spiser aldrig når den svømmer hurtigt. Slåen af cilia i mundrillen bevirker, at en kegleformet hvirvel af fødeholdigt vand bliver fejet ind i mundrillen fra en afstand forud for den forreste ende (fig. 20.16).
Næringspartiklerne går derefter til vestibulen, hvorfra nogle fødepartikler afvises og kastes ud, men andre passerer ind i cytostomet.
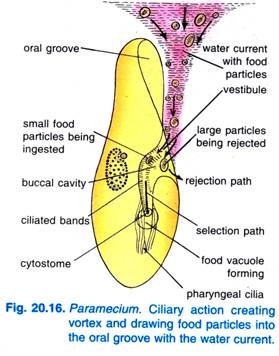
For enden af cytopharynx dannes en fødevakuole, som bliver fyldt med fødepartikler. Quadrulus og peniculi kontrollerer passagen af føden ind i fødevakuolen, som dannes lateralt. Når fødevakuolen når en vis størrelse, omslutter de postbuccale fibre fødevakuolen, og den klemmes af af dem, hvorefter den begynder at bevæge sig videre.
Den vakuole indeholder en del vand ud over fødevarerne. Roterende strømmende bevægelser af endoplasma, kaldet cykluser, fører fødevakuolerne langs et bestemt forløb, som funktionelt svarer til en fordøjelseskanal.
Kanalen begynder fra enden af cytopharynx, derefter til den bageste side, derefter fremad for at cirkulere med endoplasmaet, derefter til den dorsale overflade, derefter mod den forreste ende og derefter nedad til cytopygen. Tidligt på sin rejse mindskes fødevakuolen i størrelse, for derefter at stige igen.

Forrådnelse og ætsning hos Paramecium caudatum:
Under cyklus sker fordøjelsen ved hjælp af enzymer, der udskilles af protoplasmaet i vakuolerne. Ved fordøjelsen omdannes proteiner til aminosyrer, kulhydrater til opløselige sukkerstoffer og glykogen, og fedtstoffer fordøjes sandsynligvis også.
Indholdet af fødevakuoler er først surt (pH ca. 4) og bliver derefter alkalisk, den vigtigste fordøjelse sker i den alkaliske fase. Det ufordøjede stof egestes gennem cytopygen med en vis kraft.
Cyklose kan påvises eksperimentelt; hvis Paramecium fodres med mælk farvet med congorødt, vil mælkens fedtkugler i fødevakuolerne først blive røde på grund af enzymernes sure reaktion, derefter vil de skifte fra lilla nuancer til blå på grund af den alkaliske reaktion, vakuolerne vil vise cyklosens forløb.
Respiration og udskillelse af Paramecium Caudatum:
Gasudvekslingen (ilt og kuldioxid) foregår gennem den semipermeable pellicle ligesom hos andre ferskvandsprotozoer ved diffusionsprocessen. Paramecium Caudatum får sin ilt fra det omgivende vand. Kuldioxid og organisk affald som f.eks. ammoniak, der stammer fra stofskiftet, udskilles sandsynligvis ved at diffundere ud i vandet i den modsatte retning.

6. Osmoregulering i Paramecium Caudatum:
Paramecium Caudatum har to kontraktile vacuoler, en forreste og en bagerste. De kontraktile vacuoler har til opgave at osmoregulere, dvs. at regulere kroppens vandindhold og kan også tjene til udskillelse af kvælstofholdige affaldsstoffer som f.eks. urinstof og ammoniak.
Vandoverskud (på grund af kontinuerlig endosmosis) i cytoplasmaet udskilles i tubuli af endoplasmatisk retikulum og går til nephridial-tubuli → føderkanaler → og samles i ampulla af en serie af 6 til 11 udstrålende kanaler, der konvergerer mod og udmunder i hver vacuole. Kanalerne er mest iøjnefaldende, når en vacuole er ved at blive dannet.
Når hver vacuole er svulmet op (diastole) til en vis størrelse, trækker den sig sammen (systole) og udleder sig til ydersiden formentlig gennem en pore. De kontraktile vacuoler trækker sig skiftevis sammen med intervaller på 10-20 sekunder.
Den bageste kontraktile vacuole arbejder hurtigere end den forreste vacuole, fordi cytopharynx optager en stor mængde vand i den bageste region. De kontraktile vacuoler opretholder en optimal koncentration af vand i kroppens cytoplasma ved at bortskaffe det overskydende vand.

Adfærd hos Paramecium Caudatum:
Paramecium Caudatums reaktioner på forskellige former for stimuli lærer man ved at studere dens reaktioner og gruppering eller spredning af individer i en kultur. Reaktionen er positiv, hvis dyret bevæger sig mod en stimulus, og negativ, hvis det bevæger sig væk. På en negativ stimulus fortsætter dyret med at give den undvigende reaktion, indtil det undslipper.
I undvigelsesreaktionen vender ciliærslaget om, dyret bevæger sig et kort stykke bagud og roterer derefter i en konisk bane ved at svinge den forreste ende aboralt, mens det drejer på den bageste spids. Alle justeringer foretages ved forsøg og fejl. Forsøg har vist, at den forreste ende af dyret er mere følsom end de andre dele.
Paramecium’s reaktioner på forskellige stimuli kan grupperes som følger:
(i) Reaktioner på kontakt (Thigmotaxis):
Reaktionen på kontakt er varieret hos Paramecium. Hvis den forreste ende berøres let med en fin spids, opstår der en stærk undvigende reaktion. Når en svømmende Paramecium støder ind i en genstand i vandet, men hvis den berøres et andet sted, kan der ikke være nogen reaktion. Et langsomt bevægende individ reagerer ofte positivt på berøring med et objekt ved at komme til at hvile på det.
(ii) Reaktioner på kemikalier (kemotaksi):
Generelt reagerer Paramecia på et kemisk stimuli ved at undgå at reagere. Hvis en dråbe svag saltopløsning (0,5 %) introduceres i en Paramecium-population på en mikroglidebane, reagerer dyrene med undvigelsesreaktion, og ingen af dem kommer ind i dråben. For syrer er reaktionen imidlertid positiv, selv når koncentrationen er tilstrækkelig stærk til at dræbe dem.
(iii) Reaktioner på temperatur (Thermotaxis):
Paramecium søger en optimal temperatur på 24 til 28°C. Når en temperaturændring sker markant over eller under det optimale område, viser Paramecia en undvigende reaktion. Større varme stimulerer hurtige bevægelser og undvigende reaktioner, indtil dyrene flygter eller bliver dræbt.
(iv) Reaktioner på lys (Phototaxis):
Med undtagelse af den grønne Paramecium bursaria, som er positivt fototaktisk, er de andre arter ligeglade med almindeligt lys. Når lysintensiteten pludselig og kraftigt øges, følger der dog generelt en negativ reaktion. Paramecia udviser en umiddelbar negativ reaktion på ultraviolette stråler.
(v) Reaktioner på elektrisk strøm (Galvanotaxis):
Paramecia reagerer på elektriske stimuli. Når to elektroder placeres over for hinanden i en lavvandet skål med Paramecia og der påføres en konstant strøm, svømmer alle organismerne i samme retning mod katoden eller den negative elektrode, hvor de koncentrerer sig i stort antal.
Hvis den elektriske strøms retning vendes, mens Paramecia svømmer mod katoden, vender organismerne retningen om og svømmer mod den nye katode.

(vi) Reaktioner på vandstrøm (Rheotaxis):
Paramecia udviser en positiv rheotaxis. I en let vandstrøm bevæger Parameciaerne sig for det meste med strømmen med deres forreste ende opstrøms.
(vii) Reaktioner på tyngdekraften (Geotaxis):
Paramecia udviser generelt en negativ reaktion på tyngdekraften, som det ses i en kultur, hvor mange individer samles tæt under overfladefilmen med deres forreste ender øverst. Hvis Paramecia introduceres i et omvendt vandfyldt U-formet rør, der er stoppet i begge ender, bevæger de sig straks opad i den vandrette del af røret.
Fremstilling hos Paramecium Caudatam:
Paramecium Caudatum reproducerer sig aseksuelt ved tværgående binær fission og gennemgår også flere typer af nuklear reorganisering, såsom konjugation, endomixis, autogami, cytogami og hemixis osv.
(i) Tværgående binær fission:
Transvers binær fission er den mest almindelige form for aseksuel formering hos Paramecium. Det er en udpræget unik aseksuel proces, hvor et fuldt udvokset eksemplar deler sig i to datterindivider uden at efterlade et forældrekadaver.
Delingsplanet går gennem cellens centrum og i et plan vinkelret på kroppens længdeakse. Deling af cellelegemet som helhed går altid forud for deling af kernerne; det ser faktisk ud til, at reproduktionen indledes af kerneaktivitet og deling.

Paramecium caudatum formerer sig ved tværgående binær spaltning under gunstige forhold. Ved binær fission deler mikronukleussen sig ved mitose i to dattermikronukler, som bevæger sig til modsatte ender af cellen. Makronukleussen forlænges og deler sig på tværs ved amitose.
En anden cytopharynx knoppes af, og der opstår to nye kontraktile vacuoler, en nær den forreste ende og en anden nær den bageste ende. I mellemtiden opstår der en konstriktionsfur nær midten af kroppen og uddybes, indtil cytoplasmaet er helt delt.
De resulterende to “datterparametier” er lige store og indeholder hver et sæt celleorganeller. Af de to producerede datterparameciaer kaldes den forreste for proter og den bageste for opisthe. De vokser til fuld størrelse, før der sker en ny deling.
Den binære fissionsproces tager ca. to timer at gennemføre og kan finde sted en til fire gange om dagen, hvilket giver 2 til 16 individer. Der produceres omkring 600 generationer på et år.
Formidlingshastigheden afhænger af de ydre betingelser med hensyn til føde, temperatur, kulturens alder og befolkningstæthed; også af de indre faktorer som arvelighed og fysiologi. Naturligvis, hvis alle efterkommere af et individ skulle overleve og formere sig, ville antallet af producerede paramecia snart svare til jordens volumen.
Begrebet klon bruges til at betegne alle de individer, der er blevet produceret fra et individ ved spaltning. Alle medlemmer af en klon er arveligt ens.
(ii) Konjugation:
Ordentligt formerer Paramecium Caudatum sig ved binær fission i lange perioder, men med mellemrum kan dette afbrydes ved, at to dyr forenes langs deres mundflader med henblik på den seksuelle proces, konjugation.
Konjugation defineres som den midlertidige forening af to individer, der gensidigt udveksler mikrokernemateriale. Det er en unik type af en seksuel proces, hvor to organismer adskilles kort efter udvekslingen af kernemateriale.
Sonneborn (1947) har på grundlag af parringsadfærd hos Paramecium Caudatum rapporteret, at hver art af Paramecium findes i et antal varianter eller syngenes. Endvidere findes der inden for hver syngen et antal parringstyper normalt to.
Parringstyperne er morfologisk set identiske, men de udviser fysiologiske forskelle. Hos P. aurelia er der 14 syngenes og 28 parringstyper, mens der hos P. caudatum er 16 syngenes og 32 parringstyper. Det er blevet observeret, at paramecia normalt hverken konjugerer med medlemmer af deres egen parringstype eller med de andre sorter, men kun med den anden parringstype af deres egen sort.

Faktorer, der fremkalder konjugation:
De faktorer, der inducerer konjugation, varierer fra art til art, men nogle af dem er angivet nedenfor:
1. Konjugering sker normalt under ugunstige livsbetingelser; sult eller mangel på føde og særlig bakteriel kost eller visse kemikalier siges at fremkalde konjugeringsprocessen hos visse arter af Paramecium.
2. Konjugationen sker efter ca. 300 aseksuelle generationer af binær fission, eller den veksler med binær fission med lange mellemrum for at forynge den døende klon, dvs. den forekommer hos de individer, som skal have gennemgået et ønskeligt antal aseksuelle generationer, der siges at være perioden af umodenhed, og derefter bliver de kønsmodne til at konjugere.
3. Konjugationen sker, når der sker en ændring i parameciernes fysiologiske tilstand, så sker den mellem sådanne individer, som er noget mindre i størrelse (210 mikron lange), og de befinder sig på et stadium, der kan betragtes som en periode med usund alderdom; paramecierne i denne tilstand vil dø, hvis de ikke får lov til at konjugere.
4. Pludselig mørke i lysforhold og lave temperaturer siges at fremkalde konjugationsprocessen hos nogle arter.
5. Konjugationen finder ikke sted om natten eller i mørke; den starter tidligt om morgenen og fortsætter til om eftermiddagen.
6. Et proteinholdigt stof i cilierne hos individer af parringstypen siges at fremkalde konjugationen.
Konjugationsprocessen:
Konjugationsprocessen er forskellig hos de forskellige arter af Paramecium, men nedenstående redegørelse vedrører konjugationsprocessen hos P. caudatum (Fig. 20.21).
I konjugationen kommer to Paramecium caudatum (benævnt prækonjuganter) af modsatte parringstyper af samme art sammen med deres ventrale flader og forenes ved deres mundriller; deres cilier producerer en substans på kroppens overflade, som bevirker, at de to konjugerende paramecier klæber sammen.
De holder op med at æde, og deres mundrilleapparat forsvinder. Pelliclen og ektoplasmaet, på det sted, hvor de begge er i kontakt, går i stykker, og der dannes en protoplasmatisk bro mellem de to dyr. Nu kaldes disse individer for konjuganter.
I denne tilstand svømmer det konjugerende par aktivt, og samtidig finder der en række kerneforandringer sted i hver konjugant som beskrevet nedenfor:
Makronukleus begynder at gå i opløsning, den bliver løs i konsistensen og danner et komplekst snoet strøg, i løbet af den sidste halvdel af konjugationsperioden vil den til sidst forsvinde og blive absorberet i cytoplasmaet. Mikronukleus af hver konjugant deler sig to gange, hvoraf den ene er en reduktionsdeling.
Der produceres således fire haploide dattermikronukler i hver konjugant. Tre af disse fire mikronukler degenererer i hver, således at der kun er én tilbage.
Den resterende mikronukleus i hver konjugant deler sig mitotisk i to ulige pronuclei eller gametiske kerner, der danner en større stationær hun-prokyle og en mindre, aktiv vandrende han-prokyle.
Den ene konjugants vandrende pro-kerne krydser over den protoplasmatiske bro og smelter sammen med den anden konjugants stationære pro-kerne for at danne en synkaryon eller konjugationskerne, hvor det diploide antal kromosomer er genetableret, og hvor der er sket en udveksling af arveligt materiale.
Processen er blevet sammenlignet med befrugtning hos højere dyr, men der er ikke tale om befrugtning, fordi der ikke er nogen kønsceller involveret. Konjuganterne adskilles nu (efter ca. 12-48 timer) og kaldes ex-konjuganter. Synkaryonen i hver ex-konjugant deler sig tre gange, så der dannes otte mikronukler i hver ex-konjugant.
Fire af de otte mikronukler forstørres og bliver til makronukler, og tre af de andre fire mikronukler forsvinder.
Den resterende mikronukleus deler sig, og samtidig deler ex-konjuganten sig ved binær fission i to celler, der hver har to makronukler og en mikronukleus. Cellerne og deres mikrokerner deler sig endnu en gang for at danne fire paramecia fra hver ex-konjugant, således at de hver har en makronukleus og en mikrokyle.
Den nye makronukleus, som også mikronukleus, er blevet lavet af nyt materiale. Disse nye kerner indeholder sandsynligvis et nyt og anderledes potentiale, som afspejler sig hos de raske individer.
Betydning af konjugering:
En klon vil dø ud, hvis der ikke sker en kerneomlægning, men klonen kan forynges til at genvinde sin tidligere styrke ved kerneomlægning, denne kerneomlægning sker ved konjugering, konjugering er således afgørende for fortsat binær spaltning.
Den betydning, som konjugationen har, er opsummeret nedenfor:
1. Konjugation tjener som en foryngelses- og reorganiseringsproces, hvorved racens vitalitet genoprettes. Hvis konjugationen ikke finder sted i lange perioder, svækkes og dør parameciaerne. (Woodruffs påstand om at holde paramecia sundt i 22.000 generationer uden konjugation er modbevist af Sonneborn, fordi han viste, at alle Woodruffs paramecia tilhørte den samme parringstype).
2. Der er ingen kønsforskel hos konjuganterne, selv om kun paramecia af to forskellige parringstyper af den samme sort vil konjugere.
3. Der er ingen kønsforskel, men den aktive vandrende pro-kerne betragtes som han og den stationære pro-kerne som hun.
4. Konjugationen er kun en midlertidig forening, der sker ingen sammensmeltning af cytoplasmaet, og der dannes ingen zygote, men kernen i hver eks-konjugant indeholder arveligt materiale fra to konjugerende individer.
5. Konjugation medfører udskiftning af makronukleus med materiale fra synkaryonet, dette er en begivenhed af fundamental betydning. Ved binær fission blev makronukleusens kromosomer fordelt tilfældigt på dattercellerne, fortsat binær fission havde gjort klonen svag med nogle strukturelle abnormiteter.
Konjugation bevirker, at der dannes det korrekte antal kromosomer i makronukleus, således at racen fornyes i kraft. Mikronukleus’ rolle er at genoprette et afbalanceret kromosom- og genkompleks.
Afvigende adfærd i reproduktionen hos Paramecium Caudatum:
Paramecium Caudatum viser visse variationer i sin kerneadfærd under spaltning og konjugation, disse afvigelser er endomixis, autogami, cytogami og hemixis. Ved de tre første processer sker der genetisk rekombination, og der dannes en ny makronukleus ud fra mikronukleus.
(i) Endomixis:
Woodruff og Erdmann (1914) rapporterede først om en ny kerneomorganiseringsproces, endomixis (Gr., endon = indenfor; mixis = sammenblanding) hos Paramecium aurelia, en bimikronukleær art (Fig. 20.22). Denne proces blev beskrevet som værende periodisk forekommende, hvor der produceres et nyt makrokerneapparat uden synkaryondannelse. Endomixis forekommer inden for et enkelt individ.
I henhold til Woodruff og Erdmann degenererer makronukleussen, og mikronukleerne deler sig to gange for at danne otte mikronukler. Seks mikronukler degenererer, og to forbliver tilbage. Da der kun er to mikrokerner tilbage, deler dyret sig ved spaltning i to celler med hver en mikrokerne.
Mikronukleus i hver dattercelle deler sig to gange for at danne fire mikronukleier. To mikronukleier udvider sig til makronukleier. Dyret og dets mikronukler deler sig, således at der dannes to datterindivider med hver en makronukleus og to mikronukler.
Endomixis forekommer i den variant af P. aurelia, der ikke konjugerer, derfor kan virkningen af endomixis være den samme som konjugeringen, da begge processer medfører udskiftning af makronukleus med materiale fra mikronukleus, og begge processer forynger racens vitalitet.
Men de to processer adskiller sig fra hinanden, fordi der ikke sker nogen sammensmeltning af pronuclei i endomixis; endomixis kan sammenlignes med parthenogenese.
Men nogle arbejdere har dog med gode grunde hævdet, at endomixis ikke er en gyldig proces, og at den er blevet beskrevet på grund af fejlagtige observationer. Efter al sandsynlighed finder endomixis ikke sted, og det kan kun være et specialiseret tilfælde af autogami.

Senere rapporterede Erdmamm og Woodruff (1916) om endomixis hos Paramecium caudatum. Diller tror dog ikke på gyldigheden af denne proces og mener, at Erdmann og Woodruff simpelthen har kombineret stadierne hemixis og autogami i ét skema, endomixis.
(ii) Autogami:
Diller (1934, 1936) og Sonneborn (1950) beskrev en proces med selvbefrugtning eller autogami, der forekommer hos et enkelt individ hos Paramecium aurelia (Fig. 20.23). Han rapporterede, at ved autogami producerer tre mikrokerne (prægamiske) delinger, der indebærer modning, de gametiske kerner (pronuclei).
Ved autogami hos P. aurelia deler de to mikronukler to gange (én gang meiotisk) for at danne otte mikronukler, hvoraf seks degenererer. I mellemtiden vokser makronukleussen til en strædelignende masse, som senere brydes i stykker for at blive optaget i cytoplasmaet. To af de otte mikronukler, som pronukler, går ind i en protoplasmatisk kegle, der buler ud i nærheden af celleudmundingen.
De to pronuclei smelter sammen og danner synkaryon. Synkaryonen deler sig to gange for at danne fire mikronukler. To mikronukleier bliver til makronukleier. Parameciumet og dets mikrokerner deler sig og danner to datterindivider med hver en makronukleus og to mikrokerner. Denne proces er afsluttet på ca. to dage.
Autogami medfører en foryngelse af racen.
Det ligner konjugation for så vidt som den nye makronukleus dannes af materiale fra mikronukleus, i den nye makronukleus genetableres det korrekte antal kromosomer; og også for så vidt som der sker en sammensmeltning af to pronukler. Men autogami adskiller sig fra konjugation, fordi kun ét individ deltager i autogami, og det giver begge pronukler, det er en slags selvbefrugtning.
(iii) Hemixis:
Diller (1936) rapporterede om hemixis hos Paramecium aurelia (Fig. 20.24). Hemixis er primært en proces med makrokernefragmentering og -deling uden nogen usædvanlig mikrokerneaktivitet. Diller klassificerede hemixis i fire typer, nemlig A, B, C og D som vist i Fig. 20.24 i P. aurelia, men han stødte også på alle typer i massekulturer af P. caudatum og P. multimicronucleatum.
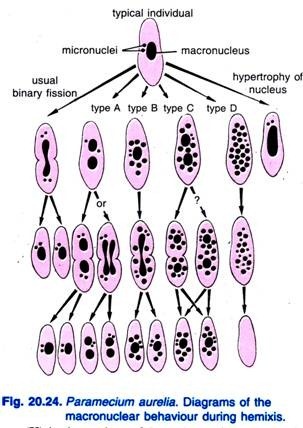
Type A er den enkleste form for hemixis, der er karakteriseret ved en opdeling af makronukleus i to eller flere dele. Denne deling er ikke synkroniseret med mikrokernedelingen.
Type B er karakteriseret ved ekstrudering af en til 20 eller flere kromatinkugler fra makronukleus til cytoplasmaet.
Type C er karakteriseret ved samtidig opsplitning af makronukleus i to eller flere større dele og ekstrudering af makronukleære kugler ud i cytoplasmaet.
Type D anses for at repræsentere patologiske tilstande, hvor makronukleus gennemgår fuldstændig fragmentering til kromatinkugler, der til sidst forsvinder fra cellen. Mikronuclei forsvinder generelt før opløsningen af makronucleus.
(iv) Cytogami:
Wichterman (1939) har rapporteret om en anden seksuel proces hos Paramecium caudatum, som han kaldte cytogami. Ved cytogami sker der ingen kerneudveksling. I denne proces kommer to individer sammen ved deres ventrale overflader, men de to individers pellicle nedbrydes ikke.
Mikronkernen i hvert individ deler sig tre gange og danner otte mikronukler, hvoraf seks af dem går i opløsning i hvert individ. De to resterende mikronukleier smelter sammen og danner en synkaryon i hver celle. Dyrene adskilles nu.
Cytogami adskiller sig fra autogami ved at der er to dyr i kontakt med hinanden, men det ligner autogami og konjugation ved sammensmeltningen af to pronuklerier. Cytogami adskiller sig fra konjugation ved, at der ikke sker nogen kerneudveksling mellem de to dyr, der kommer sammen.
Nogle cytoplasmatiske partikler rapporteret i Paramecium Caudatum:
(i) Kappa-partikler:
I 1938 rapporterede T.M. Sonneborn, at nogle racer (kendt som dræberne eller dræberstammen) af Paramecium producerer et giftigt stof, kaldet paramecin, som er dødeligt for andre individer, kaldet sensitive. Paramecinet er vandopløseligt og diffunderbart og afhænger for sin produktion af nogle partikler, der befinder sig i cytoplasmaet hos Paramecium (dræberstammen).
Disse partikler kaldes kappa-partikler. Kappa-partiklerne indeholder DNA og RNA. Et dræberparamecium kan indeholde hundredvis af kappa-partikler. Den detaljerede undersøgelse af disse partikler har afsløret, at et dominerende gen (K) i Parameciums kerne er nødvendigt for, at kappa-partiklerne kan eksistere, formere sig og producere paramecin.
(ii) mµ, Partikler:
R.W. Siegel (1952) rapporterede om en anden type dræberpartikler i cytoplasmaet hos nogle Paramecium. Et Paramecium med mµ-partikler kaldes matedræber, fordi det, når det danner konjugation med et Paramecium uden mµ-partikler, kaldet matesensitiv, dræber sidstnævnte. mp-partiklerne består også af DNA, RNA osv.
Disse partikler findes kun i de paramecia, hvis mikronukleus indeholder mindst et dominerende gen af et af de to par af ikke-forbundne kromosomale gener (M1 og M2).
(iii) Pi-partikler:
Disse partikler formodes at være den muterede form af kappa-partikler, men de producerer ikke nogen form for giftigt stof.
(iv) Lambdapartikler:
Disse partikler er rapporteret i dræberparametier og siges at producere et eller andet stof, der er ansvarligt for at forårsage lysis eller opløsning af følsomme parametier, dvs. som ikke besidder det.