Dopamiini on katekoliamiini- ja fenetyyliamiiniperheisiin kuuluva orgaaninen kemikaali. Dopamiini toimii aivojen välittäjäaineena.
Dopaminerginen signalointi liittyy palkitsemiseen motivoituneeseen käyttäytymiseen ja motoriseen kontrolliin dopamiinijärjestelmän toimintahäiriöiden johtaessa lukuisiin sairauksiin. Esimerkiksi degeneratiivinen Parkinsonin tauti johtuu dopamiinia erittävien neuronien häviämisestä, mikä johtaa motoriseen heikkenemiseen. Enzo Life Sciences tarjoaa dopamiini-ELISA-kittiä dopamiinin pitoisuuden kvantitatiiviseen mittaamiseen seerumissa, plasmassa ja soluviljelmien supernatanteissa.
Dopamiinin roolin löytäminen välittäjäaineena
Ennen vuotta 1957 vallitseva näkemys oli, että 3-hydroksityramiini oli välituotteena noradrenaliinin ja adrenaliinin synteesissä tyrosiinista. Vuosien 1957 ja 1959 välisenä aikana Kathleen Montagu ja hänen kollegansa Hans Weil-Malherben laboratoriossa Runwellin sairaalassa (Englannissa) sekä Arvin Carlsson ja hänen kollegansa Lundin yliopistossa (Ruotsissa) tekivät kuitenkin rinnakkaisia ponnisteluja, jotka auttoivat saamaan aikaan ensimmäiset havainnot, jotka yhdessä osoittivat dopamiinin roolin ihmisaivojen välittäjäaineena. Elokuussa 1957 Montagu julkaisi ensimmäisen artikkelin, joka osoitti hänen havaintonsa keskeisistä välittäjäaineista. Osana tutkimustaan hän suoritti pylväspohjaisen määrityksen, jolla tutkittiin noradrenaliinin, adrenaliinin ja 3-hydroksityramiinin määriä useiden lajien (rotta, kani, marsu, poikanen, ihminen ja sammakko) aivoista uutetuista kudoksista. Montagu arveli, että hydroksityramiinin kaltaista katekoliamiinia saattaa olla vielä yksi, jonka hän myöhemmin vahvisti olevan 3,4-dihydroksityramiinia (”dopamiinia”) paperikromatografian avulla, joka tehtiin hänen käyttämiensä uutettujen aivokudosten hartsien eluaateista. Marraskuussa 1957 Carlsson havaitsi, että hän pystyi kumoamaan akineettiset vaikutukset, joita reserpiini aiheutti hänen kaneissaan injektoimalla dopamiinin ja noradrenaliinin esiastetta, 3, 4-dihydroksifenyylialaniinia (L-DOPA) suonensisäisesti, ja havaitsi, että se korreloi dopamiinin mutta ei noradrenaliinin palautumiseen. Nämä tiedot viittasivat siihen, että dopamiinin puute saattoi olla vastuussa hänen eläimissään havaitusta akineettisesta tilasta. Lopulta Carlssonin ryhmä kehitti määrityksen, jolla voitiin mitata dopamiinipitoisuutta aivoissa, ja kartoitti, missä dopamiinin pitoisuus oli suurin. He totesivat, että dopamiinia oli suuria pitoisuuksia striatumissa, joka on tyvitumakkeiden suurin osa. Tuolloin jo tiedettiin, että tyvitumakkeilla on keskeinen rooli tahdonalaisissa motorisissa toiminnoissa. Nämä havainnot auttoivat muokkaamaan ensimmäisiä hypoteeseja, joiden mukaan dopamiini voi olla keskeinen välittäjäaine motoristen toimintojen hallinnassa.
Mitä on dopamiini? Missä sitä tuotetaan aivoissa? Miten sitä tuotetaan?
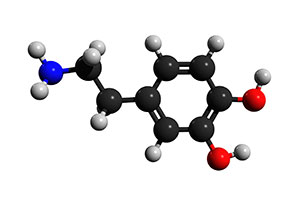
Kuva 1: Dopamiinin välittäjäainemolekyyli
Alkuperäisen löytönsä jälkeen dopamiinia, joka tunnetaan myös nimellä 3,4-dihydroksityramiini, on luonnehdittu runsaasti. Se koostuu bentseenirenkaasta, jossa on kaksi hydroksyylisivuryhmää kiinnittyneenä yhteen amiiniryhmään etyyliryhmän välityksellä. Aivojen dopaminergiset neuronit tuottavat sitä tyrosiinista lisäämällä siihen hydroksyyliryhmän, joka muuttaa sen L-DOPA:ksi (tai Levo-DOPA:ksi), ja poistamalla sen jälkeen karboksyylihapporyhmän etyylisivuketjusta, joka on kytketty amiiniryhmään, jolloin tuloksena on dopamiini. Dopaminergiset neuronit, jotka tuottavat tätä signaalimolekyyliä, sijaitsevat aivoissa substantia nigrassa ja ventraalisella tegmentaalialueella, jotka molemmat sijaitsevat keskiaivoissa, sekä hypotalamuksen kaarevassa ytimessä. Dopamiini toimii neurotransmitterina – kemikaalina, jota neuronit vapauttavat siirtääkseen sähköisen signaalin kemiallisesti hermosolujen välillä välittäen signaalin keskushermostoon ja keskushermostosta. Dopamiinin tuotannon jälkeen välittäjäaine pakataan synaptiseen vesikkeliin, vesikulaariseen monoamiinin kuljettajaan 2 (VMAT2) ja varastoidaan, kunnes toimintapotentiaalit aiheuttavat dopamiinin vapautumisen synaptiseen rakoon ja aiheuttavat sitoutumisen dopamiinireseptoreihin postsynaptisessa neuronissa.
Dopamiinivälittäjäaineet sitoutuvat viiteen dopamiinireseptorien alatyyppiin: D1, D2, D3, D4 ja D5, jotka ovat G-proteiinikytkentäisten reseptorien perheen jäseniä, jotka jakautuvat kahteen pääalaluokkaan: D-1:n kaltaiset ja D-2:n kaltaiset. Dopamiinin sitoutuminen näihin reseptoreihin käynnistää signalointikaskadit, jotka vastaavat toimintojen aktivoinnista niihin liittyvillä aivoalueilla, joilla kukin reseptorityyppi on yleisimmin läsnä. D1-tyyppiset reseptorit ovat yleisempiä kuin D2-tyyppiset reseptorit. Sen ymmärtäminen, miten dopamiini toimii ihmisaivoissa välittäjäaineena, edellyttää, että tarkastellaan dopamiinin sitoutumista D1:n kaltaisiin ja D2:n kaltaisiin reseptorityyppeihin, jotta se voisi vaikuttaa toisten sanansaattajajärjestelmien kautta. Dopamiinin sitoutuminen D1:n kaltaisiin reseptoreihin (D1 ja D5) johtaa eksitaatioon Na+-kanavien avautumisen kautta tai inhibitioon K+-kanavien avautumisen kautta. D1:n kaltaisen reseptorin stimulaatio indusoi adenylaattisyklaasin, entsyymin, joka muuntaa ATP:n cAMP:ksi, aktivoitumisen, jolloin cAMP-tasot nousevat, mikä johtaa proteiinikinaasi A:n (PKA), joka fosforyloi alempana olevia kohteita, kuten cAMP:n säätelevää elementtiä sitovaa proteiinia (CREB). CREB:n siirtyminen ytimeen ja CREB:stä riippuva lukuisten geenien transkriptio ovat vastuussa oppimisen ja muistin muodostumisen edellyttämästä synaptisesta plastisuudesta. Sitä vastoin D-2:n kaltaisten reseptorien (D2, D3 ja D4) sitoutuminen johtaa kohdehermosolun estämiseen, sillä ne vaikuttavat päinvastoin estämällä adenylaattisyklaasia kytkeytymällä G-proteiinien Gi/o:han, mikä vähentää cAMP:n tuotantoa. Se, onko dopamiini kiihottavaa vai estävää, riippuu siitä, minkä tyyppinen vaikutus kohdehermosoluun kohdistuu, mikä perustuu siihen, minkä tyyppisiä reseptoreita hermosolun kalvopinnalla on ja miten hermosolu reagoi cAMP-pitoisuuden nousuun tai laskuun.
Mitä dopamiini tekee ihmisen aivoissa?
Dopamiinilla on tärkeitä rooleja toimeenpanevissa toiminnoissa, motorisessa kontrollissa, motivaatiossa, kiihottumisessa, vahvistumisessa ja palkitsemisessa signaalikaskadien kautta, jotka vaikuttavat sitoutumalla dopaminergisiin reseptoreihin ihmisaivojen substantia nigrassa, ventraalisella tegmentaalialueella ja hypotalamuksen kaarevassa ytimessä olevissa projektioissa.
Substantia nigrassa nigro-striataalinen rata projisoi dopaminergisiä neuroneja tuloalueelta (joka tunnetaan nimellä pars compacta) dorsaaliseen striatumiin, ja sillä on ensisijainen rooli motoristen toimintojen hallinnassa ja motoristen taitojen oppimisessa. Jos nigro-striataalisen radan dopaminergiset neuronit rappeutuvat, tämä aiheuttaa motorisen kontrollin häiriöitä, mikä on Parkinsonin taudin tunnusmerkki.
Ventraalisella tegmentaalisella alueella (VTA) mesolimbinen rata projisoituu prefrontaalisesta aivokuoresta amygdalan ytimeen (nucleus accumbens), cingulaariseen gyrusiin, hippokampukseen ja hajulohkon pyriformiseen kompleksiin. Amygdalan ja cingulaarisen gyrusin dopaminergiset projektiot ovat vastuussa tunteiden muodostumisesta ja käsittelystä. Hippokampuksessa dopaminergisten neuronien läsnäolo liittyy oppimiseen, työmuistiin ja pitkäkestoisen muistin muodostumiseen. Lopuksi hajulohkon pyriforminen kompleksi on vastuussa siitä, että ihmiset saavat hajuaistin. Mesolimbisessä reitissä dopamiinia vapautuu nautinnollisten tilanteiden aikana, mikä aiheuttaa kiihtymystä ja vaikuttaa käyttäytymiseen (motivaatioihin), jotta etsitään nautinnollista toimintaa tai ammattia, ja se sitoutuu ytimessä accumbensissa ja prefrontaalisessa aivokuoressa oleviin dopaminergisiin reseptoreihin. Nucleus accumbensiin suuntautuvien projektioiden lisääntyneellä aktiivisuudella on suuri merkitys vahvistumisessa ja äärimmäisissä tapauksissa riippuvuuksissa.
Hypotalamuksen kaarevassa ytimessä dopamiinineuronit muodostavat tuberoinfundibulaarisen radan, joka projisoituu aivolisäkkeeseen ja estää prolaktiinihormonin eritystä. Arcuate nucleus -ytimen neuronien tuottamaa dopamiinia vapautuu hypotalamo-hypofyysisiin verisuoniin, jotka toimittavat aivolisäkkeeseen dopamiinia prolaktiinin tuotannon estämiseksi.
Dopamiinitasojen kvantitatiivinen mittaus näytteistä
Dopamiinitasojen arvioiminen siltä osin kuin ne liittyvät tiettyihin aivojen osiin liittyvien toimintojen säätelyn heikkenemiseen, on houkutteleva kohde neurotieteelliselle tutkimukselle. Enzo Life Sciences tarjoaa Dopamine ELISA Kitin, joka on kolorimetrinen kilpailullinen immunomääritys, jolla voidaan kvantifioida dopamiinia seerumissa, plasmassa, kudoshomogenaateissa ja muissa biologisissa nesteissä. Tämä kitti on erittäin spesifinen ihmisen dopamiinille, ja sen ristireaktiivisuus ihmisen dopamiinin ja sen analogien havaitsemisen välillä on vähäinen. Tämän erittäin herkän immunomäärityksen havaitsemisalue on niinkin pieni kuin 1,56 ng/ml ja niin suuri kuin 100 ng/ml (kuva 1). Lisäksi tämä tuote on suunniteltu niin, että sillä on korkea herkkyys, korkea eräkohtainen toistettavuus ja alhainen tuloksiin kuluva aika, ja se tarjoaa yksinkertaisen protokollan, jonka avulla loppukäyttäjille voidaan tuottaa luotettavia, kvantitatiivisia tuloksia alle kahdessa tunnissa jopa 40 näytteestä kahtena kappaleena.
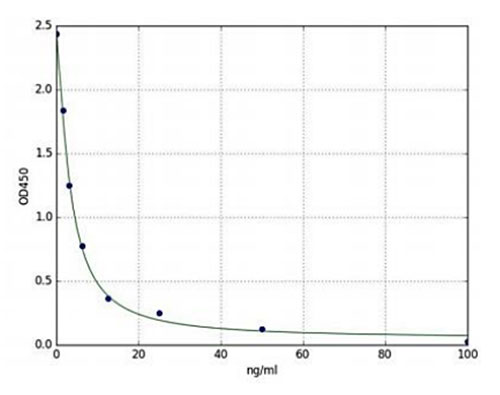
Kuva 2: Dopamiini ELISA Kitin (ENZ-KIT188) standardikäyrä, joka kuvaa tyypillistä standardikäyrää (1,56 ng/ml-100ng/ml).
Enzo Life Sciences tarjoaa laajan valikoiman tuotteita neurotieteen ja immunologian tutkimustarpeisiin. Tarjoamme muita neurotransmitterien immunomäärityksiä, kuten Serotoniini ELISA Kit ja Histamiini ELISA Kit, sekä laajan valikoiman vasta-aineita neurotransmittereiden, kuten ACTH:n, ANP:n, BNP:n, CCK:n, CGRP:n, NPY:n, GABA:n, GLP-1:n ja Substance P:n, tutkimiseen. Enzon kattavaan tuotevalikoimaan kuuluu myös SCREEN-WELL Neurotransmitter Library -kirjastomme, joka sisältää 661 CND-reseptoriligandia 96-kuopan muodossa. Katso lisätietoja Neurotieteiden ja Solusignaloinnin/signaalinsiirron alustoistamme tai ota rohkeasti yhteyttä tekniseen tukipalveluumme saadaksesi lisäapua.