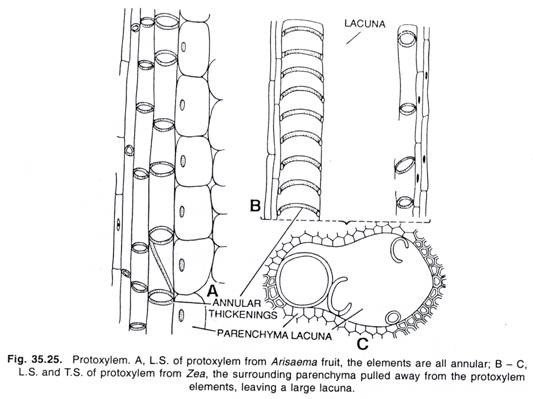
LÄHTEET:
Tässä artikkelissa keskustelemme Paramecium Caudatumista:- 1. Paramecium Caudatumin elinympäristö, elinympäristö ja kulttuuri 2. Paramecium Caudatum. Paramecium Caudatumin rakenne 3. Liikkuminen 4. Ravitsemus 5. Hengitys ja erittyminen 6. Osmoregulaatio 7. Käyttäytyminen 8. Lisääntyminen 9. Poikkeava käyttäytyminen lisääntymisessä 10. Eräät sytoplasmahiukkaset.
Sisältö:
- Habit, Paramecium Caudatumin elinympäristö ja viljely
- Paramecium Caudatumin rakenne
- Paramecium Caudatumin liikkuminen
- Paramecium Caudatumin ravitsemus
- Paramecium Caudatumin hengitys ja erittyminen
- Paramecium Caudatumin hapetus ja elimistön tasapaino. Paramecium Caudatum
- Behaviour of Paramecium Caudatum
- Reproduction in Paramecium Caudatum
- Aberrant Behaviour in Reproduction in Paramecium Caudatum
- Some Cytoplasmic Particles Reported in Paramecium Caudatum
- Habit, Paramecium Caudatumin elinympäristö ja viljely:
- Parameciumin viljely:
- Paramecium caudatumin rakenne:
- (i) Paramecium caudatumin koko ja muoto:
- (ii) Nahka:
- (iii) Cilia:
- (iv) Infra-Ciliary System:
- (v) Suuaukko ja sytopyge:
- (vi) Sytoplasma:
- (vii) Ektoplasma:
- (viii) Trikokystat:
- (ix) Neuromotorium ja siihen liittyvät fibriilit:
- (x) Endoplasma:
- (xi) Supistuvat vacuolit:
- (xii) Ravintovakuolit:
- Paramecium Caudatumin liikkuminen:
- (i) Metabolia eli vartalon vääristymät:
- (ii) Ciliary Locomotion:
- Paramecium Caudatumin ravitsemus:
- Ruokintamekanismi:
- Paramecium Caudatumin ruoansulatus ja nieleminen:
- Paramecium Caudatumin hengitys ja erittyminen:
- Paramecium Caudatumin käyttäytyminen:
- (i) Reaktiot kosketukseen (Thigmotaxis):
- (ii) Reaktiot kemikaaleihin (kemotaksis):
- (iii) Reaktiot lämpötilaan (termotaksis):
- (iv) Reaktiot valoon (fototaksis):
- (v) Reaktiot sähkövirtaan (galvanotaksis):
- (vi) Reaktiot vesivirtaan (reotaksis):
- (vii) Reaktiot painovoimaan (geotaksis):
- Reproduktio Paramecium Caudatamilla:
- (i) Poikittainen binäärinen jakautuminen:
- (ii) Konjugaatio:
- Paramecium Caudatumin lisääntymiskäyttäytyminen:
- (i) Endomiksis:
- (ii) Autogamia:
- (iii) Hemixis:
- (iv) Sytogamia:
- Joitakin Paramecium Caudatumissa raportoituja sytoplasmahiukkasia:
- (i) Kappa-hiukkaset:
- (ii) mµ, hiukkaset:
- (iii) Pi-partikkelit:
- (iv) Lambda-hiukkaset:
Habit, Paramecium Caudatumin elinympäristö ja viljely:
Paramecium caudatum (Gr., paramekes = pitkulainen; L., caudata = pyrstö) esiintyy yleisesti makean veden lammikoissa, altaissa, ojissa, puroissa, järvissä, tekoaltaissa ja joissa. Sitä esiintyy erityisesti runsaasti seisovissa lammissa, joissa on runsaasti hajoavaa ainesta, orgaanisissa infuusioissa ja jätevesissä. Paramecium caudatum on vapaasti elävä organismi, ja tämä laji on levinnyt maailmanlaajuisesti.
Parameciumin viljely:
VAROITUKSET:
Viedään lammesta upotettuja rikkaruohoja ja asetetaan tislatulla vedellä täytettyyn purkkiin, peitetään purkki ja jätetään se mätänemään; Paramecia-parvia ilmestyy muutamassa päivässä. Keitä nyt heinää vedessä, dekantoi infuusio ja lisää muutama vehnänjyvä ja anna sen seistä, kunnes se on sameaa bakteereista.
Siirrä Paramecia ensimmäisestä purkista tähän nesteeseen, jossa ne lisääntyvät nopeasti. Pelkästään heinäinfuusio tuottaa Parameciaa, joka osoittaa kystien läsnäoloa, ja hiekanjyviä muistuttavia kystia on raportoitu, mutta ei ole todisteita siitä, että Paramecia muodostaa kystia, koska niitä ei ole koskaan vahvistettu.
Paramecium caudatumin rakenne:
(i) Paramecium caudatumin koko ja muoto:
Paramecium caudatum (kuva 20.1) on mikroskooppinen organismi ja paljain silmin havaittavissa pienenä pitkänomaisena kappaleena.
HAVAINNOT:
Se näyttää vaaleanharmaalta tai valkoiselta ja mittaa tavallisesti 170-290 mikronia ja voi saavuttaa jopa 300-350 mikronin pituuden. P. caudatum näyttää tossun tai kengänpohjan näköiseltä, minkä vuoksi eläintä kutsutaan yleisesti tossueläimeksi. Se on neljä kertaa niin pitkä kuin leveä ja jokseenkin lieriömäinen, ja sen päät ovat selvästi erilaiset.
Eteenpäin liikkuva etuosa on hoikka ja sen pää on tylppä tai pyöristetty, kun taas takaosa on hieman terävä tai kartiomainen. Elimistön levein osa on hieman keskikohdan alapuolella. Eläimen ruumis on muodoltaan epäsymmetrinen, ja siinä on selvästi erottuva suu- tai ventraalipinta ja aboraalipinta tai selkäpinta.
(ii) Nahka:
Vartaloa peittää ohut, kaksikerroksinen, elastinen ja luja, gelatiinista valmistettu pellikli. Pellilli pitää eläimen muodon, mutta on riittävän joustava salliakseen supistukset. Pellikkelissä on kaksoiskalvo, josta ulompi kalvo on yhtenäinen värekarvojen kanssa ja sisempi kalvo ektoplasman kanssa. Mikroskoopin suuremmassa suurennoksessa pellikkelin pinnalla on suorakulmaisia tai kuusikulmaisia painaumia.

Tämä järjestely on säilynyt Parameciumin selkäpinnalla, mutta ventraalipinnalla harjanteet lähenevät toisiaan edestä ja takaa preoraalista ja postoraalista aukkoa kohti. Kussakin kuusikulmaisessa syvennyksessä on keskellä aukko, jonka läpi tulee ulos yksittäinen cilium. Kuusikulmaisten syvennysten etu- ja takareunoilla on trikokystien aukot.
Ehretin ja Powersin (1957) elektronimikroskooppinen tutkimus pellikkelistä (kuva 20.2) on paljastanut, että kuusikulmaiset syvennykset vastaavat säännöllisiä ontelosarjoja, alveoleja. Kaikki alveolit muodostavat yhdessä yhtenäisen alveolikerroksen, jota rajaavat ulompi alveolikalvo ja sisempi alveolikalvo.
Uloin kerros on tiiviissä kosketuksessa ulomman solukalvon alla. Pellikkeliin kuuluu siis ulompi solukalvo, ulompi alveolikalvo ja sisempi alveolikalvo.
(iii) Cilia:
Koko elimistöä peittävät lukuisat, pienet, karvamaiset ulokkeet, joita kutsutaan värekarvoiksi. Cilia esiintyy pituussuuntaisina riveinä koko kehossa, tämä tila tunnetaan holotrichous-tilana, jossa kehon cilia on yhtä suuri. Hiukset ovat rakenteeltaan samanlaisia kuin flagellat, niillä on ulompi protoplasmavaippa tai plasmakalvo, jossa on yhdeksän kaksoispitkittäistä pituussuuntaista fibrilliä perifeerisen renkaan sisällä. Joissakin säikeissä yhdeksän ulompaa fibrilliä ei ole pareittain.
HAVAINNOT:
Keskellä on kaksi pitkittäistä fibrilliä, jotka ovat ohuempia kuin ulommat fibrillit. Kukin cilium syntyy basaalisesta granulasta tai kinetosomista. Yhdeksän paria perifeerisiä fibrillejä sulautuu yhteen muodostaen kinetosomin seinämän, joten kinetosomi on putki, joka on joko avoin tai suljettu alapäässään, kaksi keskimmäistä fibrilliä pysähtyvät useimmissa sädekehissä pellikkelin tasolle.
Kinetosomista lähtee ohut rihmasto, joka ei liity ytimeen. Monilla Metazooilla on myös sädekehiä, niiden rakenne on sama, paitsi että tyvirake on erilainen ja siinä on hienoja säikeitä tai juurisäikeitä, jotka ulottuvat alas sytoplasmaan. Hiukset eroavat kuitenkin flagelloista siinä, että niitä on yleensä enemmän ja ne ovat kooltaan lyhyempiä.
Hiilarit voidaan kätevästi jakaa vartalo- eli somaattisiin hiusläikkiin, joita on vartalon pinnalla, ja suun alueen hiusläikkiin, jotka liittyvät suun alueeseen. Vartalohiukset ovat samanmuotoisia, mutta ne ovat pidempiä takapäässä, mistä johtuu nimi caudatum. Hiukset ovat liikkumisen ja ravinnon keräämisen elimiä, ja ne toimivat myös aistinvaraisina reseptoreina ja havaitsevat ulkoisen ympäristön ärsykkeet.

Hiilion rakenne:
Hiukset ja lippulat ovat koostumukseltaan fibrillaarisia. Ciliumin halkaisija on tyvestä noin 0,2 mikronia tai 2000 A0, joka voi olla jopa 10 mikronia solun pinnan yläpuolella. Hiuskalvoja rajoittaa 90 A0:n paksuinen yksikkökalvo, joka muistuttaa plasmakalvoa ja on jatkuva plasmakalvon kanssa. Ciliumin rajattu tila sisältää vesipitoisen aineen, jota kutsutaan matriisiksi.
Matriisiin jää upotettuna yksitoista pitkittäistä fibrilliä eli mikrotubulusta. Yhdestätoista fibrillistä kaksi sijaitsee keskellä, kun taas loput yhdeksän fibrilliä jäävät sijoittumaan perifeerisesti keskeisten fibrillien ympärille. Kukin yhdeksästä ulommasta fibrillistä on halkaisijaltaan 360A0 ja koostuu kahdesta alafibrillistä, joiden halkaisija on 180-250A0.
Näitä alafibrillejä nimitetään alafibrilli A:ksi ja alafibrilli B:ksi.
HAVAINNOT:
Subfibrilli A on hieman suurempi kuin subfibrilli B. Subfibrilli A antaa toiselta puoleltaan kaksi paksua uloketta tai käsivartta. Kaikkien uloimpien fibrillien subfibrilli A:n varret pysyvät suunnattuina myötäpäivään. Lisäksi alafibrilli A sijaitsee lähempänä ciliumin keskustaa kuin alafibrilli B. Molemmilla alafibrilleillä on yhteinen seinämä, jonka paksuus on 50A0.
Kahdessa keskimmäisessä fibrillissä ei ole parittaisia subfibrillejä kuten yhdeksässä perifeerisessä fibrillissä, vaan kussakin on vain yksi putkilo. Kummankin keskusfibrillin halkaisija on noin 250A0 ja se koostuu 60A0 paksusta seinämästä.
Kummankin keskifibrillin välissä on 350A0 tilaa, ja ne pysyvät yhteisen vaipan ympäröiminä. Gibbnos (1967) on raportoinut, että keskusfibrillien tuppi antaa yhdeksän säteittäisesti suuntautunutta linkkiä tai kehää kullekin alafibrillille A.

Korkearesoluutioinen elektronimikroskooppi on paljastanut, että kukin cilia- ja flagella- eli lippalankojen periferia- ja keskusfibrilli koostuu kymmenestä tai kahdestatoista filamenttisäikeestä (filamenttisäikeestä), jotka ovat paksuudeltaan 40A0. Kukin filamentti on helmimäinen. Kukin helmi pysyy järjestäytyneenä 40 kertaa 50A0:n kokoisiin ristikoihin tubuluksen seinämän tasossa. Näitä helmiä pidetään tubuluksen rakenteen perusyksikkönä.
(iv) Infra-Ciliary System:
Infra-ciliaarinen järjestelmä sijaitsee heti pellikulaaristen alveolien alapuolella. Se koostuu kinetosomista eli tyvirungosta ja kinetodesomista. Hiukset lähtevät kinetosomeista, ja jokaisesta kinetosomista lähtee hienojakoinen sytoplasmafibrilli, jota kutsutaan kinetodesmaksi, (kuva 20.2).
Pelliksen alapuolella hieman oikealle, mutta yhdistyneenä kaikkiin yhden pitkittäisrivin kinetosomeihin, on useiden kinetodesomien pituussuuntainen nippu, jokaisen kinetosomin kinetodesoma ulottuu jonkin matkan verran etupuolelle omaan kinetodesmojen nippuunsa.
Kinetosomien pituussuuntainen rivi kinetodesmatoineen muodostaa pitkittäisen yksikön, jota kutsutaan kinetoksi. Kaikki kinetiat tai kinetiat muodostavat säilykeläisen infrakiliaalijärjestelmän. Kinetiat sijaitsevat kuoressa pellikkelin alapuolella, ja niiden määrä on melko vakio jokaisessa säilykelajissa.
Infraciliaarinen järjestelmä ohjaa ja koordinoi värekarvojen liikkeitä, ja se saa aikaan organellien muodostumisen solunjakautumisessa, esim. jotkut kinetiat muodostavat suun. Ciliattien binäärisessä jakautumisessa kinetiat leikkautuvat poikittain kahtia, joista kumpikin menee yhteen tytärsoluun, tätä kutsutaan perikinetaaliseksi jakautumiseksi.
(v) Suuaukko ja sytopyge:
Ventrolateraalisella puolella on suuri vino, matala syvennys, jota kutsutaan suu-uraksi tai peristomaksi ja joka antaa eläimelle epäsymmetrisen ulkonäön. Se kulkee vinosti toiselta puolelta taaksepäin (yleensä vasemmalta oikealle, mutta joissakin tapauksissa oikealta vasemmalle) ja päättyy hieman keskirungon taakse. Suuaukko johtaa lyhyeen kartiomaisen suppilon muotoiseen syvennykseen, jota kutsutaan eteiseksi.
HAVAINNOT:
Vestibuli johtaa suoraan kiinteään, soikean muotoiseen aukkoon, jota kutsutaan sytostomiksi (suu). Sytostomista suoraan kohti kehon keskiosaa ulottuu leveä sytofarynx. Sytofarynx kääntyy sitten jyrkästi kohti takapuolta ja muuttuu hoikaksi, kapenevaksi ruokatorveksi.
Esofagus on siis suunnilleen samansuuntainen Parameciumin ruumiinpinnan kanssa lukuun ottamatta sen takaosaa. Täällä ruokatorvi kääntyy jälleen kohti eläimen keskiosaa johtaakseen muodostuvaan ravintotyhjiöön.
Sytopyykki (jota kutsutaan myös solun peräaukoksi tai peräaukon pisteeksi tai sytoproktiksi) sijaitsee rungon ventraalipinnalla lähes pystysuorassa sytostomin eli suun takana. Sytopygen kautta poistuvat sulamattomat ravintohiukkaset. Sytofarynxin solunulkoinen rakenne on hyvin monimutkainen.
Gelei (1934) ilmoitti neljän rivin olemassaolosta ja Lund (1941) havaitsi ainakin neljä riviä. Sytofarynxin vasemmassa seinämässä on rakenne nimeltä penniculus, joka kiertyy spiraalimaisesti noin 90 astetta niin, että sen takimmainen ääripää on ruokatorven suun puoleisella (ventraalisella) pinnalla.
Lundin mukaan penniculus koostuu kahdeksasta rivistä värekarvoja, jotka on järjestetty kahteen lähekkäin olevaan lohkoon. Samanlaista kaistaa, joka koostuu neljästä rivistä pitkiä värekarvoja, jotka ovat vähemmän tiiviitä kuin penniculuksessa, kutsutaan quadrulukseksi. Se kiertyy spiraalimaisesti pitkin suuontelon dorsaaliseinämää ja päättyy lähelle penniculusta.
Jotkut työntekijät ovat virheellisesti kutsuneet penniculusta ja quadrulusta aaltoilevaksi kalvoksi. Quadrulus ja penniculus kontrolloivat ravinnon kulkua. Ei tiedetä, miten värekarvat toimivat, luultavasti niiden fibrillit supistuvat rytmikkäästi, mikä aiheuttaa taivutusta. Gelei (1925) huomautti, että penniculuksen tehtävänä on ruoka-aineiden pakottaminen elimistöön.

(vi) Sytoplasma:
Sytoplasma erottuu kapeaan, ulkoiseen eli kortikaaliseen vyöhykkeeseen, jota kutsutaan ektoplasmaksi, ja laajempaan, sisäiseen eli medullaariseen alueeseen, jota kutsutaan endoplasmaksi.
(vii) Ektoplasma:
Ektoplasma (ectosarc tai cortex) on elimistön pysyvä osa, joka erottuu silmiinpistävästi endoplasmasta. Ektoplasma muodostaa kiinteän, kirkkaan, ohuen ja tiheän ulkokerroksen. Se sisältää trikokystat, värekarvoja ja fibrillaarisia rakenteita, ja sitä rajoittaa ulkoisesti pelliksi kutsuttu peite.
(viii) Trikokystat:
Ektoplasmassa suorassa kulmassa pintaan nähden on pieniä karanmuotoisia pusseja, joita kutsutaan trikokystoiksi. Pieni täplä kuusikulmion kummallakin etu- ja takareunalla merkitsee trikokystan sijaintia. Ne on täytetty taittuvalla, tiheällä nesteellä, jossa on turvottavaa ainetta, ja niiden ulommassa päässä on kartiomainen pää tai piikki.
Trikokystat sijaitsevat kohtisuorassa ektoplasmassa, ne avautuvat pienistä huokosista pellikkelin kuusikulmaisten alueiden harjanteilla.
Ne syntyvät värekarvojen kinetosomeista, vaeltavat sitten ja asettuvat samalle etäisyydelle endoplasmassa. Kun eläintä ärsytetään, trikokystat purkautuvat pitkinä tahmeina säikeinä. Purkautuneessa trikokystassa on läpinäkymätön piikki, joka muistuttaa käänteistä naulaa, ja pitkä raidallinen varsi, mutta varsi ei näy purkautumattomassa tilassa, ja se muodostuu todennäköisesti purkautumisen aikana.
Trikokystien tehtävästä ei ole varmuutta, mutta ne purkautuvat reaktiona paikallisiin kontakteihin ja vammoihin, ja ne saattavat toimia puolustusorganelleina.
Mutta tämä on epävarmaa, koska trikokystat ovat tehottomia Didiniumia vastaan, joka on Parameciumin tärkein saalistaja, ne voivat olla eläimen kiinnittämistä johonkin paikkaan ruokinnan aikana. Joissakin sädesienissä trikokystat toimivat hyökkäysorganelleina. Kun trikokystat poistuvat, ne regeneroituvat kinetosomeista.


(ix) Neuromotorium ja siihen liittyvät fibriilit:
Lundin (1933) mukaan sytofarynxin vasemmalla dorsaalisella seinämällä noin sytostomin takareunan korkeudella on hyvin pieni, kaksilohkoinen massa, neuromotorium. Neuromotoriumista säteilevät fibrillit endoplasmaan.
Näistä neljä tai useampi kulkee tavallisesti lähes selkärangan seinämään asti, mutta loput ovat lyhyempiä, eikä niiden sijainti ole selvä. Kaikkia kutsutaan endoplasmisiksi fibrilleiksi. Niiden funktio on tuntematon, mutta ne saattavat koordinoida suulakiharojen syöttöliikkeitä. Fibrillit voivat myös antaa mekaanista tukea, elastisuutta, supistuvuutta, johtavuutta ja aineenvaihdunnallista vaikutusta.

(x) Endoplasma:
Endoplasma eli medulla on sytoplasman nestemäisempi ja tilavampi osa, joka sisältää monia sytoplasmallisia granuloita sekä muita sulkeumia ja luonteeltaan erikoistuneita rakenteita. Sytoplasman sulkeumia ovat mm. mitokondriot, Golgin apparaatit, vakuolit, kiteet, rakeet ja kromidiat. Endoplasmassa on myös muita rakenteita, kuten ytimiä, supistuvia vakuoleja ja ravintovakuoleja.
Tumat:
Endoplasmassa lähellä sytostomia on kaksi ydintä, eli Paramecium on heterokaryoottinen, suuri ellipsinmuotoinen ja rakeinen makrotumake ja toinen pieni kompakti mikrotumake.
Makrotydin on silmiinpistävä, ellipsin tai munuaisen muotoinen kappale. Se on kompakti tyyppi, joka sisältää hienoja säikeitä ja tiiviisti pakkautuneita, vaihtelevan kokoisia diskreettejä kromatiinirakeita, jotka on upotettu akromaattiseen matriisiin. Siinä on paljon nukleoleja ja paljon enemmän kromatiinimateriaalia (DNA).
Se on somaattinen eli kasvullinen tuma. Se jakautuu amitoottisesti ja ohjaa eläimen vegetatiivisia toimintoja (aineenvaihduntaa). Se käy läpi mitoosin.
Mikroydin on pieni, tiivis ja pallomainen. Se sijaitsee yleensä lähellä makronukleusta usein koverana. Hienot kromatiinirakeet ja -langat ovat jakautuneet tasaisesti koko rakenteeseen. Mikrotuma jakautuu mitoottisesti ja ohjaa lisääntymistä.
Mikrotumakkeiden lukumäärä vaihtelee lajeittain; P. caudatumissa niitä on yksi, P. aureliassa kaksi ja P. multimicronucleatumissa monta. P. aureliassa mikrotumakkeessa on selvä nukleoli, mutta P. caudatumissa sitä ei ole. Moses (1949; 1950) on raportoinut, että makronukleus ja mikronukleus ovat kemialliselta koostumukseltaan identtisiä.
(xi) Supistuvat vacuolit:
Tässä on kaksi suurta, nestetäytteistä supistumiskykyistä vakuolia, joista kumpikin sijaitsee lähellä rungon toista päätä lähellä selkäpintaa. Niiden sijainti on kiinteä (toisin kuin ameeboilla), ne sijaitsevat ektoplasman ja endoplasman välissä, mutta ne ovat tilapäisiä organelleja, jotka katoavat ajoittain. Joissakin lajeissa niillä näyttää olevan limakalvo, jolloin ne eivät katoa kokonaan systolen aikana.
Kunkin supistuvan tyhjiön yhteydessä on viidestä kahteentoista putkimaista sädekanavaa, joista kukin koostuu pääteosasta, pitkästä ampullasta, joka tyhjentyessään luhistuu, ja lyhyestä injektorikanavasta, joka avautuu tyhjiöön.

Kanavat ovat yhteydessä suureen osaan elimistöä, josta ne ottavat nesteitä ja vuodattavat ne vakuoliin, joka näin muodostuu uudelleen ja kasvaa kooltaan, kun supistuva vakuoli saavuttaa maksimikokonsa, se supistuu äkillisesti (systole) ja purkaa sisältönsä pellikkelissä olevan pysyvän huokosen kautta, minkä jälkeen kanavat muodostavat jälleen supistuvan vakuolin, kanavat eivät katoa kokonaan, koska ne ovat pysyviä rakenteita.
Kaksi supistuvaa vakuolia purkautuvat epäsäännöllisesti, takimmainen supistuu nopeammin, koska se on lähellä sytofarynxiä ja siihen tulee enemmän vettä. Kanavien ja supistuvien vacuolien päätehtävä on hydrostaattinen, ne poistavat ylimääräisen veden protoplasmasta, vesi imeytyy ja imeytyy osittain ravinnon aikana.
Typpeä sisältävässä jätemateriaalissa on ammoniakkiyhdisteitä ja jonkin verran uraatteja, jotka erittyvät supistuvista vakuoleista yhdessä hiilidioksidin kanssa, mutta ei ole todisteita siitä, että protoplasma erittäisi erittyvää ainetta kanaviin tai supistuviin vakuoleihin.
Sytofarynxin toisella puolella on peristomin alapuolella kiinteä pysyvä sytopyykki eli peräaukko, jonka kautta sulamattomat ruoan jäännökset ja uraatit poistuvat.
Elektronimikroskooppinen tutkimus supistuvista vakuoleista on paljastanut, että kukin supistuva apparaatti koostuu joistakin endoplasmisen retikulumin tubuluksista, nephridiaalisista tubuluksista, syöttökanavista, lisävakuoleista (säteittäisistä kanavista) ja supistuvasta päävakuoleista. Aksessoristen vacuolien oletetaan olevan syöttökanavien ampulleja (kuva 20.11).
(xii) Ravintovakuolit:
Nämä ovat endoplasmassa sijaitsevia, suunnilleen pallonmuotoisia, kooltaan ja lukumäärältään vaihtelevia supistumattomia elimiä. Ne sisältävät nieltyjä ravintohiukkasia, pääasiassa bakteereja, ja pienen määrän nestettä, jota rajoittaa ohut, selkeä kalvo. Volkonsky (1934) ehdotti näille vakuoleille nimitystä gastriolit. Ruokavakuoleihin liittyvät ruoansulatusrakeet.
Paramecium Caudatumin liikkuminen:
Paramecium Caudatum suorittaa liikkumisen kahdella menetelmällä, nimittäin metabolialla eli vartalon vääntelyllä ja värekarvojen avulla.
(i) Metabolia eli vartalon vääristymät:
Paramecium Caudatumin vartalolla on elastisuutta, se voi puristaa itsensä vartaloaan kapeamman käytävän läpi, minkä jälkeen vartalo ottaa normaalin muotonsa. Tämä ruumiin muodon tilapäinen muutos on metabolia, sen saa aikaan Parameciumissa protoplasma.
(ii) Ciliary Locomotion:
Hiilien aikaansaama lokomootio on tärkein menetelmä. Hiukset voivat lyödä eteen- tai taaksepäin, jolloin eläin voi uida etu- tai takaperin.
Normaalisti eläin ui eteenpäin, värekarvojen lyödessä taaksepäin, mutta vinosti, värekarvat jäykistyvät ja taipuvat nopeasti taaksepäin melkein koskettamaan ruumiin pintaa, tätä kutsutaan tehoiskuksi; sen jälkeen värekarvat veltostuvat ja palaavat hitaasti alkuperäiseen pystysuoraan asentoon, tätä kutsutaan , recovery stroke.
Saman poikittaisrivin säikeet lyövät yhdessä ja saman pitkittäisrivin säikeet lyövät peräkkäin etupäästä takapäähän.

Tätä koordinoitua värekarvojen liikettä kutsutaan metakronaaliseksi rytmiksi, joka johtuu infra-ciliaarijärjestelmästä; tämä saa eläimen uimaan eteenpäin. Mutta kun vartalon värekarvat lyövät viistosti taaksepäin, niin samaan aikaan suuontelon pidemmät värekarvat lyövät voimakkaammin, mikä saa etupään kääntymään vasemmalle.
Rungon ja suuontelon värekarvojen toiminta saa eläimen pyörimään pitkän akselinsa ympäri. Tämä kierto on aina vasemmalle (paitsi P. calkinsi, joka pyörii oikeanpuoleisessa spiraalissa).
Tämä eteenpäin suuntautuvan liikkeen, väistöliikkeen ja pyörimisen yhdistelmä saa eläimen liikkumaan eteenpäin vastapäivään spiraalimaisesti. Tällä radalla on suora akseli, ja eläimen sama ruumiinpinta pysyy kohti spiraaliradan akselia. Mutta uidessaan taaksepäin kaikki lajit pyörivät oikealle.
Säiliöiden syke voidaan kääntää päinvastaiseksi niin, että säikeet liikkuvat vinosti eteenpäin, jolloin eläin ui taaksepäin. Paramecium liikkuu säikeiden avulla nopeudella, joka on 1500 mikronia tai jopa enemmän sekunnissa.
Jennings väitti, että Parameciumin spiraalimainen liikerata johtuu siitä, että vaikka värekarvat iskevät pääasiassa taaksepäin, ne iskevät viistosti oikealle aiheuttaen siten eläimen kääntymisen vasemmalle.
Tämä vartalon kääntyminen kohti aboraalista pintaa johtuu suurelta osin myös siitä, että suulakiharojen, jotka iskevät suoremmin taaksepäin, tehokas isku on voimakkaampi. Tulos – Parameciumin kiertyminen pitkän akselinsa ympäri – mahdollistaa sen, että Paramecium voi seurata enemmän tai vähemmän suoraa rataa muodostaessaan suuria spiraaleja.
Paramecium Caudatumin ravitsemus:
Paramecium Caudatumissa ravitsemus on holotsoottista. Ravinto koostuu pääasiassa bakteereista ja pienistä alkueläimistä. Paramecium ei odota ravintoa vaan metsästää sitä aktiivisesti.
Väitetään, että Paramecium Caudatum osoittaa valinnanvaraa ravintonsa valinnassa, mutta tälle ei näytä olevan perusteita, vaikka se nielaisee vain tietyntyyppisiä bakteereja; käytettävissä olevien tietojen mukaan yksi Paramecium ahmii 2-5 miljoonaa Bacillus coli -yksilöä 24 tunnissa. Se syö myös yksisoluisia kasveja, kuten leviä ja piileviä, sekä pieniä eläin- ja vihannespaloja.
Ruokintamekanismi:
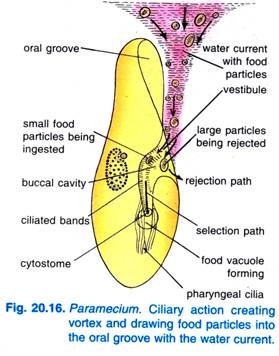
Kun Paramecium Caudatum tulee alueelle, jossa on runsaasti ravintoa, se tulee lepäämään. Se syö vain levossa tai uidessaan hyvin hitaasti, se ei koskaan syö uidessaan nopeasti. Suuontelon värekarvojen lyöminen saa aikaan sen, että ruokaa sisältävästä vedestä muodostuu kartiomainen pyörre, joka pyyhkäistään suuonteloon etäämmältä etupäästä (kuva 20.16).
Ruokahiukkaset kulkeutuvat sitten eteiseen, josta osa ruokahiukkasista hylätään ja heitetään ulos, mutta osa kulkeutuu sytostomiin.
Sytofarynxin päähän muodostuu ravintotyhjiö, joka täyttyy ruokahiukkasilla. Quadrulus ja peniculi ohjaavat ruoan kulkua ravintotyhjiöön, joka muodostuu sivusuunnassa. Kun ravintotyhjiö saavuttaa tietyn koon, post-buccal-kuidut tarttuvat ravintotyhjiöön, ja ne puristavat sen irti, jolloin se lähtee liikkeelle.
Ruokavakooli sisältää ravinnon lisäksi jonkin verran vettä. Endoplasman pyörivät virtaavat liikkeet, joita kutsutaan sykloosiksi, kuljettavat ravintovakuoleja tiettyä rataa pitkin, joka vastaa toiminnallisesti ruoansulatuskanavaa.
Reitti alkaa sytofarynxin päästä, sitten takapuolelle, sitten eteenpäin kiertämään endoplasman mukana, sitten selkäpinnalle, sitten kohti etupäätä, sitten alaspäin sytopygeen. Matkansa alkuvaiheessa ravintotyhjiön koko pienenee ja kasvaa sitten uudelleen.

Paramecium Caudatumin ruoansulatus ja nieleminen:
Sykloosin aikana ruoansulatus tapahtuu protoplasmasta vakuoleihin erittyvien entsyymien avulla. Ruuansulatuksessa proteiinit muuttuvat aminohapoiksi, hiilihydraatit liukoisiksi sokereiksi ja glykogeeniksi, ja todennäköisesti myös rasvat pilkkoutuvat.
Ruokavakuolien sisältö on aluksi hapanta (pH noin 4) ja muuttuu sitten emäksiseksi, pääasiallinen ruoansulatus tapahtuu emäksisen vaiheen aikana. Sulamaton aines egestoituu sytopyykin läpi jonkin verran voimaa käyttäen.
Sykloosi voidaan osoittaa kokeellisesti; jos Parameciumille syötetään Kongon punaisella värjättyä maitoa, ravintovakuolissa olevat maidon rasvapallot muuttuvat ensin punaisiksi entsyymien happaman reaktion vuoksi, sitten ne muuttuvat violetin sävyistä sinisiksi emäksisen reaktion vuoksi, vakuolit osoittavat sykloosin kulun.
Paramecium Caudatumin hengitys ja erittyminen:
Kaasujen (happi ja hiilidioksidi) vaihto tapahtuu muiden makean veden alkueläinten tapaan diffuusion avulla puoliläpäisevän pellikkelin läpi. Paramecium Caudatum saa hapensa ympäröivästä vedestä. Hiilidioksidi ja aineenvaihdunnasta peräisin olevat orgaaniset jätteet, kuten ammoniakki, erittyvät todennäköisesti diffundoitumalla ulospäin veteen päinvastaiseen suuntaan.

6. Osmoregulaatio Paramecium Caudatumissa:
Paramecium Caudatumissa on kaksi supistuvaa vakuolia, toinen etu- ja toinen takapuolella. Supistuvien vacuolien tehtävänä on osmoregulaatio eli elimistön vesipitoisuuden säätely, ja ne voivat toimia myös typpipitoisten jätteiden, kuten urean ja ammoniakin, erittymisessä.
Sytoplasmassa oleva ylimääräinen vesi (jatkuvan endosmoosin vuoksi) erittyy endoplasmisen retikulumin tubuluksiin ja kulkeutuu nephridiaalisiin tubuluksiin → syöttökanaviin → ja kerääntyy ampulliin, joka koostuu 6-11 säteittäisen kanavan sarjasta, jotka lähenevät toisiaan kohti ja purkautuvat kuhunkin vakuoliin. Kanavat ovat selvimmin havaittavissa tyhjiön muodostuessa.
Kun kukin vakuoli on paisunut (diastole) tiettyyn kokoon, se supistuu (systole) ja purkautuu ulospäin todennäköisesti huokosen kautta. Supistuvat vakuolit supistuvat vuorotellen, 10-20 sekunnin välein.
Takimmainen supistuva vakuoli toimii nopeammin kuin etummainen vakuoli, koska sytofarynx imee suuren määrän vettä takimmaiseen alueeseen. Supistuvat vakuolit pitävät yllä optimaalista vesipitoisuutta kehon sytoplasmassa hävittämällä ylimääräisen veden.

Paramecium Caudatumin käyttäytyminen:
Paramecium Caudatumin reaktiot erilaisiin ärsykkeisiin opitaan tutkimalla sen reaktioita ja yksilöiden ryhmittymistä tai hajaantumista viljelyssä. Reaktio on positiivinen, jos eläin liikkuu kohti ärsykettä, ja negatiivinen, jos se liikkuu poispäin. Epäsuotuisaan ärsykkeeseen eläin jatkaa välttävää reaktiota, kunnes se pakenee.
Välttämisreaktiossa sädekehän lyönti kääntyy päinvastaiseksi, eläin liikkuu lyhyen matkan taaksepäin ja pyörii sitten kartiomaisella radalla heilauttamalla etupäätä aboraalisesti samalla kun se kääntyy takakärjen varassa. Kaikki säädöt tehdään kokeilemalla ja erehtymällä. Kokeet ovat osoittaneet, että eläimen etupää on herkempi kuin muut osat.
Parameciumien reaktiot eri ärsykkeisiin voidaan ryhmitellä seuraavasti:
(i) Reaktiot kosketukseen (Thigmotaxis):
Vasteet kosketukseen vaihtelevat Parameciumissa. Jos etupäätä kosketetaan kevyesti hienolla kärjellä, syntyy voimakas välttämisreaktio. Kun uiva Paramecium törmää johonkin vedessä olevaan esineeseen, mutta jos sitä kosketetaan muualla, reaktiota ei välttämättä esiinny. Hitaasti liikkuva yksilö reagoi usein positiivisesti kosketukseen jonkin esineen kanssa tulemalla lepäämään sen päälle.
(ii) Reaktiot kemikaaleihin (kemotaksis):
Yleensä Paramecia reagoi kemialliseen ärsykkeeseen välttämällä reaktiota. Jos mikroliuskalla olevaan Paramecium-populaatioon tuodaan pisara heikkoa suolaliuosta (0,5 prosenttia), eläimet reagoivat välttämisreaktiolla, eikä yksikään eläimistä mene pisaraan. Happoihin reaktio on kuitenkin positiivinen, vaikka konsentraatio olisi riittävän voimakas tappamaan ne.
(iii) Reaktiot lämpötilaan (termotaksis):
Paramecium hakeutuu optimaaliseen 24-28 °C:n lämpötilaan. Kun lämpötilan muutos tapahtuu selvästi optimaalisen alueen ylä- tai alapuolella, Paramecia osoittaa välttävää reaktiota. Suurempi kuumuus saa eläimet liikkumaan nopeasti ja välttelemään reaktioita, kunnes ne pakenevat tai kuolevat.
(iv) Reaktiot valoon (fototaksis):
Vihreää Paramecium bursariaa lukuun ottamatta, joka on positiivisesti fototaksinen, muut lajit suhtautuvat tavalliseen valoon välinpitämättömästi. Kun valon voimakkuutta kuitenkin lisätään äkillisesti ja jyrkästi, seuraa yleensä negatiivinen reaktio. Parameciat reagoivat välittömästi negatiivisesti ultraviolettisäteisiin.
(v) Reaktiot sähkövirtaan (galvanotaksis):
Paramecia reagoi sähköärsykkeisiin. Kun kaksi elektrodia asetetaan vastakkain matalaan Parameciaa sisältävään astiaan ja siihen kytketään vakiovirta, kaikki organismit uivat samaan suuntaan kohti katodia eli negatiivista elektrodia, johon ne keskittyvät suurina määrinä.
Jos sähkövirran suunta käännetään, kun Paramecia ui kohti katodia, organismit kääntävät suunnan ja uivat kohti uutta katodia.

(vi) Reaktiot vesivirtaan (reotaksis):
Paramecia osoittaa positiivista reotaksista. Lievässä vesivirtauksessa Paramecia liikkuu enimmäkseen virtauksen mukana etupäätään ylävirtaan.
(vii) Reaktiot painovoimaan (geotaksis):
Paramecia reagoi yleensä negatiivisesti painovoimaan, mikä näkyy viljelyssä, jossa monet yksilöt kerääntyvät tiiviisti pintakalvon alle etupäät ylhäällä. Jos Paramecia viedään ylösalaisin olevaan, vedellä täytettyyn U-muotoiseen putkeen, joka on suljettu molemmista päistä, ne siirtyvät välittömästi ylöspäin putken vaakasuoraan osaan.
Reproduktio Paramecium Caudatamilla:
Paramecium Caudatum lisääntyy suvuttomasti poikittaisella binäärisellä jakautumisella ja käy läpi myös useita erilaisia ydinten uudelleenorganisoitumisen muotoja, kuten konjugaatio, endomixis, autogamia, sytogamia ja hemixis jne.
(i) Poikittainen binäärinen jakautuminen:
Transversaalinen binäärinen fissio on yleisin suvuttoman lisääntymisen tyyppi Parameciumissa. Se on selvästi ainutlaatuinen suvuton prosessi, jossa yksi täysikasvuinen yksilö jakautuu kahdeksi tytäryksilöksi jättämättä vanhempien ruumista.
Jakaantumistaso kulkee solun keskipisteen läpi ja tasossa, joka on suorassa kulmassa rungon pituusakseliin nähden. Koko solurungon jakautumista edeltää aina ytimien jakautuminen; näyttääkin siltä, että lisääntyminen käynnistyy ydintoiminnasta ja jakautumisesta.

Paramecium caudatum lisääntyy suotuisissa olosuhteissa poikittaisella binäärisellä jakautumisella. Binäärisessä fissiossa mikrotuma jakautuu mitoosin avulla kahdeksi tytärmikrotumaksi, jotka siirtyvät solun vastakkaisiin päihin. Makrotydin pidentyy ja jakautuu poikittain amitoosin avulla.
Toinen sytofaryyni irtoaa ja kaksi uutta supistuvaa vakuolia ilmestyy, toinen lähellä etupäätä ja toinen lähellä takapäätä. Samaan aikaan supistumiskouru ilmestyy lähelle rungon keskiosaa ja syvenee, kunnes sytoplasma on kokonaan jakautunut.
Tuloksena syntyvät kaksi ”tytär” parameciaa ovat samankokoisia, ja kumpikin sisältää joukon soluelimiä. Kahdesta syntyvästä tytärparaméciasta etummaista kutsutaan proteriksi ja takimmaista opistiksi. Ne kasvavat täyteen kokoonsa ennen uutta jakautumista.
Kaksoistumisprosessi kestää noin kaksi tuntia, ja se voi tapahtua yhdestä neljään kertaa päivässä, jolloin syntyy 2-16 yksilöä. Vuodessa syntyy noin 600 sukupolvea.
Monistumisnopeus riippuu ulkoisista olosuhteista, kuten ravinnosta, lämpötilasta, kulttuurin iästä ja populaatiotiheydestä, sekä sisäisistä tekijöistä, kuten perinnöllisyydestä ja fysiologiasta. Luonnollisesti, jos yhden yksilön kaikki jälkeläiset jäisivät henkiin ja lisääntyisivät, tuotettujen paramecioiden määrä vastaisi pian maapallon tilavuutta.
Käsitteellä klooni tarkoitetaan kaikkia niitä yksilöitä, jotka on tuotettu yhdestä yksilöstä jakautumalla. Kaikki kloonin jäsenet ovat perinnöllisesti samanlaisia.
(ii) Konjugaatio:
Ordinomaisesti Paramecium Caudatum lisääntyy binäärisen jakautumisen avulla pitkiä aikoja, mutta ajoittain tämä voi keskeytyä kahden yksilön liittyessä yhteen suupintojaan pitkin suvulliseen konjugaatioprosessiin.
Konjugaatio määritellään kahden sellaisen yksilön väliaikaiseksi liitoksi, jotka vaihtavat keskenään mikroydinmateriaalia. Se on ainutlaatuinen seksuaalisen prosessin tyyppi, jossa kaksi organismia eroaa toisistaan pian ydinaineen vaihdon jälkeen.
Sonneborn (1947) on Paramecium Caudatum -lajin parittelukäyttäytymisen perusteella ilmoittanut, että kullakin Paramecium-lajilla on useita lajikkeita eli syngeenejä. Lisäksi kunkin syngenin sisällä on useita parittelutyyppejä, joita on yleensä kaksi.
Parittelutyypit ovat morfologisesti identtisiä, mutta niissä on fysiologisia eroja. P. aureliassa on 14 syngeeniä ja 28 parittelutyyppiä, kun taas P. caudatumissa on 16 syngeeniä ja 32 parittelutyyppiä. On tehty havaintoja siitä, että yleensä parameciat eivät konjugoi oman parittelutyyppinsä jäsenten eivätkä muiden lajikkeiden kanssa, vaan ainoastaan oman lajikkeensa toisen parittelutyypin kanssa.

Konjugaatiota aiheuttavat tekijät:
Konjugaatiota aiheuttavat tekijät vaihtelevat lajeittain, mutta joitakin niistä on esitetty alla:
1. Konjugaatio tapahtuu yleensä epäsuotuisissa elinolosuhteissa; nälänhädän tai ravinnon puutteen ja tietyn bakteeriruokavalion tai tiettyjen kemikaalien sanotaan aiheuttavan konjugaatioprosessin tietyissä Paramecium-lajeissa.
2. Konjugaatio tapahtuu noin 300 suvuttoman sukupolven binäärisen jakautumisen jälkeen, tai se vuorottelee binäärisen jakautumisen kanssa pitkin väliajoin kuolevan kloonin nuorentamiseksi, eli se tapahtuu yksilöissä, joiden on täytynyt läpäistä toivottava määrä suvuttomia sukupolvia, joiden sanotaan olevan epäkypsyyskausi, minkä jälkeen niistä tulee sukupuolisesti kypsiä, jotta ne voivat konjugoitua.
3. Konjugoituminen tapahtuu, kun paramecien fysiologisessa tilassa tapahtuu muutos, silloin se tapahtuu sellaisten yksilöiden välillä, jotka ovat kooltaan jonkin verran pienempiä (210 mikronin pituisia) ja ne ovat vaiheessa, jota voidaan pitää epäterveellisen vanhuuden kautena; tämän tilan paramecia kuolee, jos sen ei anneta konjugoitua.
4. Äkillisen pimeyden valo-olosuhteissa ja matalien lämpötilojen sanotaan aiheuttavan konjugaatioprosessin joissakin lajeissa.
5. Konjugaatio ei tapahdu yöllä tai pimeässä; se alkaa varhain aamulla ja jatkuu iltapäivään asti.
6. Parittelutyypin yksilöiden värekarvoissa olevan valkuaisaineen sanotaan aiheuttavan konjugaatiota.
Konjugaatioprosessi:
Konjugaatioprosessi vaihtelee eri Paramecium-lajeissa, mutta alla oleva selostus koskee P. caudatum -lajin konjugaatioprosessia (kuva 20.21).
Konjugaatiossa kaksi saman lajin vastakkaisia parittelutyyppejä edustavaa Paramecium caudatum -parameciumia (joita kutsutaan prekonjugaatioiksi) lähestyvät toisiaan ventraalipinnoiltaan ja yhdistyvät suuonteloidensa välityksellä; niiden värekarvat tuottavat rungon pinnalle ainetta, joka saa aikaan kahden konjugoituvan parameciumin tarttumisen.
Ne lopettavat syömisen ja niiden suuontelolaite häviää. Molempien pellikyyli ja ektoplasma hajoavat kosketuskohdassa, ja näiden kahden eläimen välille muodostuu protoplasmaattinen silta. Nyt näitä yksilöitä kutsutaan konjuganteiksi.
Tässä tilassa konjugoituva pari ui aktiivisesti, ja samanaikaisesti kummassakin konjugantissa tapahtuu sarja jäljempänä kuvattuja ydinmuutoksia:
Makrotydin alkaa hajota, se muuttuu rakenteeltaan löysäksi ja muodostaa monimutkaisen kieroutuneen sukkulan, konjugaatioajan jälkimmäisen puoliskon aikana se lopulta häviää imeytyen sytoplasmaan. Kunkin konjugantin mikrotuma jakautuu kahdesti, joista toinen on pelkistymisjakautuma.
Siten jokaisessa konjugantissa syntyy neljä haploidista tytärmikrotydintä. Kolme näistä neljästä mikrotumasta degeneroituu kummassakin, niin että jäljelle jää vain yksi.
Kunkin konjugantin jäljelle jäävä mikrotumake jakautuu mitoottisesti kahdeksi eriarvoiseksi pronukleukseksi tai gameettiseksi ytimeksi, jotka muodostavat suuremman paikallaan pysyvän naaraspuolisen pro-nukleuksen ja pienemmän, aktiivisen, vaeltavaisen urospuolisen pro-nukleuksen.
Toisen konjugantin vaeltava pro-ydin ylittää protoplasmasillan ja sulautuu toisen konjugantin paikallaan pysyvän pro-ytimen kanssa muodostaen synkaryonin tai konjugaatioytimen, jossa kromosomien diploidinen lukumäärä on palautunut ja jossa perintöaines on vaihtunut.
Prosessia on verrattu korkeampien eläinten hedelmöitymiseen, mutta kyseessä ei ole hedelmöityminen, koska siihen ei liity sukusoluja. Konjugantit eroavat nyt (noin 12-48 tunnin kuluttua) toisistaan ja niitä kutsutaan ex-konjuganteiksi. Kunkin ex-konjugantin synkaryoni jakautuu kolme kertaa muodostaen kussakin ex-konjugantissa kahdeksan mikrotumia.
Kahdeksasta mikrotumasta neljä suurenee ja muuttuu makrotumiksi, ja kolme neljästä muusta mikrotumasta häviää.
Jäljelle jäävä mikrotuma jakautuu ja samalla ex-konjugantti jakautuu binäärisen jakautumisen kautta kahdeksi soluksi, joissa kummassakin on kaksi makrotumaa ja yksi mikrotuma. Solut ja niiden mikrotumat jakautuvat toisen kerran muodostaen kustakin ex-konjugantista neljä parameciaa siten, että kussakin on yksi makrotuma ja yksi mikrotuma.
Uusi makrotydin, kuten myös mikrotydin, on tehty uudesta materiaalista. Nämä uudet ytimet sisältävät todennäköisesti uutta ja erilaista potentiaalia, joka näkyy terveissä yksilöissä.
Konjugaation merkitys:
Klooni kuolee sukupuuttoon, jos ytimen uudelleenjärjestäytymistä ei tapahdu, mutta klooni voidaan nuorentaa palauttamaan entinen elinvoimansa ytimen uudelleenjärjestäytymisellä, tämä ytimen uudelleenjärjestäytyminen saadaan aikaan konjugaatiolla, joten konjugaatio on välttämätön binäärisen fission jatkumisen kannalta.
Konjugaation merkitys on tiivistetty alla:
1. Konjugaatio toimii nuorentumis- ja uudelleenorganisoitumisprosessina, jonka avulla rodun elinvoima palautuu. Jos konjugaatiota ei tapahdu pitkiä aikoja, paramecia heikentyy ja kuolee. (Woodruffin väitteen, jonka mukaan hän oli pitänyt parameciat terveinä 22 000 sukupolven ajan ilman konjugaatiota, kumosi Sonneborn, koska hän osoitti, että kaikki Woodruffin parameciat kuuluivat samaan pariutumistyyppiin).
2. Konjuganttien sukupuolta ei eroteta toisistaan, vaikka konjugoituvat vain saman lajikkeen kahta eri pariutumistyyppiä edustavat parameciat.
3. Sukupuolta ei eroteta toisistaan, mutta aktiivista vaeltavaa pro-nukleusta pidetään kuitenkin uroksena ja paikallaan olevaa pro-nukleusta naaraana.
4. Konjugoituminen on vain väliaikainen liitto, sytoplasmaa ei fuusioidu eikä zygoottia synny, mutta kunkin entisen konjugantin ydin sisältää perintöainesta kahdesta konjugoituvasta yksilöstä.
5. Konjugaatio saa aikaan makrotumakkeen korvautumisen synkaryonista peräisin olevalla materiaalilla, tämä on perustavanlaatuisen tärkeä tapahtuma. Binaarisessa fissiossa makronukleuksen kromosomit jakautuivat sattumanvaraisesti tytärsoluille, jatkuva binäärinen fissio oli tehnyt kloonista heikon, jossa oli joitakin rakenteellisia poikkeavuuksia.
Konjugaatio saa aikaan oikean kromosomimäärän muodostumisen makrotumakkeeseen, jolloin rotu uudistuu elinvoimaisena. Mikrotumakkeen tehtävänä on palauttaa tasapainoinen kromosomi- ja geenikompleksi.
Paramecium Caudatumin lisääntymiskäyttäytyminen:
Paramecium Caudatum osoittaa tiettyjä poikkeamia ydinkäyttäytymisessä fission ja konjugaation aikana, nämä poikkeamat ovat endomixis, autogamia, sytogamia ja hemixis. Kolmessa ensimmäisessä prosessissa tapahtuu geneettinen rekombinaatio ja mikrotumasta muodostuu uusi makrotumake.
(i) Endomiksis:
Woodruff ja Erdmann (1914) raportoivat ensimmäisenä uudesta ytimen uudelleenorganisoitumisprosessista, endomixiksestä (gr., endon = sisällä; mixis = sekoittuminen) Paramecium aurelia -lajissa, joka on bimikronukleaattinen laji (kuva 20.22). Tämän prosessin kuvattiin tapahtuvan määräajoin, jolloin syntyy uusi makroydinlaite ilman synkaryonin muodostumista. Endomixis esiintyy yhden yksilön sisällä.
Woodruffin ja Erdmannin mukaan makronukleus degeneroituu ja mikronukleukset jakautuvat kahdesti muodostaen kahdeksan mikronukleusta. Kuusi mikrotumia degeneroituu ja kaksi jää jäljelle. Kun jäljelle jää vain kaksi mikrotumia, eläin jakautuu jakautumalla kahdeksi soluksi, joissa kummassakin on yksi mikrotuma.
Kummankin tytärsolun mikrotuma jakautuu kahdesti muodostaen neljä mikrotumaa. Kaksi mikrotumia suurenee muodostaen makrotumia. Eläin ja sen mikrotumat jakautuvat siten, että muodostuu kaksi tytäryksilöä, joissa kummassakin on yksi makrotuma ja kaksi mikrotumaa.
Endomiksaatiota esiintyy siinä P. aurelia -lajikkeessa, joka ei konjugoi, joten endomiksaation vaikutus voi olla sama kuin konjugaation, koska molemmat prosessit saavat aikaan makrotumakkeen korvautumisen mikrotumakkeesta peräisin olevalla materiaalilla ja molemmat prosessit nuorentavat rodun elinvoimaa.
Mutta nämä kaksi prosessia eroavat toisistaan, koska endomiksaatiossa ei tapahdu pronukleoiden fuusiota; endomiksaatiota voidaan verrata parthenogeneesiin.
Jotkut tutkijat ovat kuitenkin perustellusti väittäneet, että endomiksis ei ole pätevä prosessi ja että se on kuvattu virheellisen havainnoinnin vuoksi. Todennäköisesti endomiksistä ei tapahdu, ja se voi olla vain autogamian erikoistapaus.

Myöhemmin Erdmamm ja Woodruff (1916) raportoivat endomiksaatiosta Paramecium caudatumilla. Diller ei kuitenkaan usko tämän prosessin paikkansapitävyyteen ja katsoo, että Erdmann ja Woodruff ovat yksinkertaisesti yhdistäneet hemixis- ja autogamian vaiheet yhdeksi järjestelmäksi, endomixikseksi.
(ii) Autogamia:
Diller (1934, 1936) ja Sonneborn (1950) kuvasivat Paramecium aureliassa (kuva 20.23) yksittäisessä yksilössä esiintyvän itsehedelmöitymisprosessin eli autogamian. Hän kertoi, että autogamiassa kolme mikroydinjakautumaa (pregamiajakautumaa), joihin liittyy kypsyminen, tuottavat gameettiset ytimet (pronuclei).
P. aurelian autogamiassa kaksi mikrotumaketta jakautuu kahdesti (kerran meioottisesti) muodostaen kahdeksan mikrotumaketta, joista kuusi degeneroituu. Samaan aikaan makronukleus kasvaa sukkulamaiseksi massaksi, joka myöhemmin hajoaa palasiksi ja imeytyy sytoplasmaan. Kahdeksasta mikrotumasta kaksi siirtyy pronukleuksina protoplasmakartioon, joka pullistuu lähelle solun suuta.
Kaksi pronuklea sulautuvat synkaryoniksi. Synkaryoni jakautuu kahdesti muodostaen neljä mikrotumia. Kahdesta mikrotumasta tulee makrotumia. Paramecium ja sen mikrotumat jakautuvat muodostaen kaksi tytäryksilöä, joissa kummassakin on yksi makrotuma ja kaksi mikrotumaa. Tämä prosessi päättyy noin kahdessa päivässä.
Autogamia saa aikaan rodun nuorentumisen.
Se muistuttaa konjugaatiota sikäli, että uusi makrotumake muodostuu mikrotumakkeesta peräisin olevasta materiaalista, uudessa makrotumakkeessa palautuu oikea kromosomimäärä; ja myös siinä, että tapahtuu kahden pronukleuksen fuusio. Autogamia eroaa kuitenkin konjugaatiosta, koska vain yksi yksilö osallistuu autogamiaan, ja se tuottaa molemmat pronomyytit, se on eräänlainen itsensä hedelmöittäminen.
(iii) Hemixis:
Diller (1936) raportoi hemixisistä Paramecium aureliassa (kuva 20.24). Hemixis on ensisijaisesti makrotumallisten ydinten pirstoutumis- ja jakautumisprosessi ilman epätavallista mikrotumallisten ydinten toimintaa. Diller luokitteli hemiaksin neljään tyyppiin, nimittäin A, B, C ja D, kuten kuvassa 20.24 on esitetty P. aureliassa, mutta hän havaitsi kaikkia tyyppejä myös P. caudatumin ja P. multimicronucleatumin massaviljelmissä.
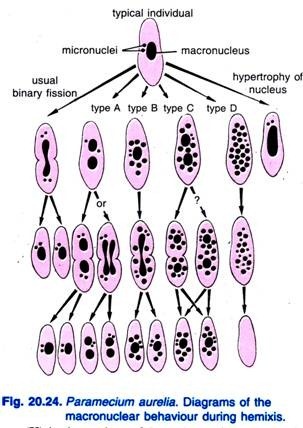
Tyyppi A on yksinkertaisin hemixis-muoto, jolle on ominaista makronukleuksen jakautuminen kahteen tai useampaan osaan. Tämä jakautuminen ei ole synkronoitu mikroydinjakautumisen kanssa.
Tyyppi B:lle on ominaista yhden-20 tai useamman kromatiinipallon purkautuminen makrotumakkeesta sytoplasmaan.
Tyypille C on ominaista makrotumakkeen samanaikainen jakautuminen kahteen tai useampaan suureen osaan ja makrotumakepallojen purkautuminen sytoplasmaan.
Tyypin D katsotaan edustavan patologisia tiloja, joissa makrotydin pirstoutuu kokonaan kromatiinipalloiksi, jotka lopulta häviävät solusta. Mikrotumat häviävät yleensä ennen makrotuman hajoamista.
(iv) Sytogamia:
Wichterman (1939) on raportoinut toisesta Paramecium caudatumissa esiintyvästä seksuaalisesta prosessista, jota hän kutsui sytogamiaksi. Sytogamiassa ei tapahdu ydinvaihtoa. Tässä prosessissa kaksi yksilöä tulee yhteen ventraalipinnoiltaan, mutta näiden kahden yksilön pellikki ei hajoa.
Kunkin yksilön mikrosydän jakautuu kolmesti muodostaen kahdeksan mikrosydäntä, joista kuusi hajoaa kussakin yksilössä. Kaksi jäljelle jäävää mikrotumaketta sulautuvat ja muodostavat synkaryonin kussakin solussa. Eläimet eroavat nyt toisistaan.
Sytogamia eroaa autogamiasta siinä, että kaksi eläintä on kosketuksissa toisiinsa, mutta se muistuttaa autogamiaa ja konjugaatiota siinä, että kaksi pronuklea sulautuu. Sytogamia eroaa konjugaatiosta siinä, että kahden yhteen tulevan eläimen välillä ei tapahdu ydinvaihtoa.
Joitakin Paramecium Caudatumissa raportoituja sytoplasmahiukkasia:
(i) Kappa-hiukkaset:
Vuonna 1938 T.M. Sonneborn raportoi, että jotkin Paramecium-lajin rodut (jotka tunnetaan tappajina tai tappajakantana) tuottavat paramecin-nimistä myrkyllistä ainetta, joka on tappava muille yksilöille, joita kutsutaan herkiksi. Paramekiini on vesiliukoinen ja diffuusiokykyinen, ja sen tuotanto riippuu joistakin Parameciumin (tappajakannan) sytoplasmassa olevista hiukkasista.
Näitä hiukkasia kutsutaan kappa-hiukkasiksi. Kappa-partikkeleissa on DNA:ta ja RNA:ta. Tappava Paramecium voi sisältää satoja kappa-partikkeleita. Näiden hiukkasten yksityiskohtainen tutkimus on paljastanut, että Parameciumin ytimessä oleva dominoiva geeni (K) on välttämätön, jotta kappa-hiukkaset voivat olla olemassa, lisääntyä ja tuottaa paramesiinia.
(ii) mµ, hiukkaset:
R.W. Siegel (1952) on raportoinut toisenlaisista killerihiukkasista joidenkin Parameciumien sytoplasmassa. Parameciumia, jossa on mµ-hiukkasia, kutsutaan mate killeriksi, koska kun se konjugoituu Parameciumiin, jossa ei ole mµ-hiukkasia, joita kutsutaan mate sensitive -hiukkasiksi, niin se tappaa jälkimmäisen. mp-hiukkaset koostuvat myös DNA:sta, RNA:sta jne.
Nämä hiukkaset ovat olemassa vain niissä paramecioissa, joiden mikrotuma sisältää vähintään yhden dominoivan geenin jommastakummasta kahdesta yhdistämättömästä kromosomigeeniparista (M1 ja M2).
(iii) Pi-partikkelit:
Näiden hiukkasten oletetaan olevan kappa-hiukkasten mutanttimuoto, mutta ne eivät tuota minkäänlaista myrkyllistä ainetta.
(iv) Lambda-hiukkaset:
Näiden hiukkasten on raportoitu esiintyvän tappajaparamekaaneissa, ja niiden sanotaan tuottavan jotakin ainetta, joka on vastuussa sellaisten herkkien paramekaanien lyysin tai hajoamisen aiheuttamisesta, joilla sitä ei siis ole.