Sukusolut ja hedelmöittyminen yleiskatsaus
Seksuaalinen lisääntyminen tarkoittaa vanhempien tuottamia sukupuolisia sukusoluja (sukusoluja), joista tulee biseksuaalisten sukusolujen (kuten siittiöiden ja munasolujen) yhdistelmällä hedelmöittynyt munasolu, jonka jälkeen hedelmöittyneestä munasolusta muodostuu uusi yksilö, jota kutsutaan seksuaalisuudeksi lisääntymiseksi. Sukupuolisessa lisääntymisessä tapahtuva geenien yhdistelmien laaja vaihtelu voi lisätä jälkeläisten kykyä sopeutua luonnonvalintaan. Seksuaalisen lisääntymisen jälkeläisissä sattumanvaraisesti yhdistyvät geenit voivat olla lajin kannalta hyödyllisiä tai sitten eivät, mutta ainakin ne lisäävät muutamien yksilöiden selviytymismahdollisuuksia arvaamattomassa ja jatkuvasti muuttuvassa ympäristössä, mikä hyödyttää lajia. Sukupuolinen lisääntyminen voi myös edistää suotuisten mutaatioiden leviämistä populaatiossa. Jos lajin kahdella yksilöllä on suotuisia mutaatioita eri paikoissa, suvuttomassa lisääntymispopulaatiossa nämä kaksi mutanttia kilpailevat keskenään, kunnes yksi eliminoituu, molempien suotuisten mutaatioiden samanaikainen säilyminen on mahdotonta. Suvullisesti lisääntyvissä populaatioissa nämä kaksi suotuisaa mutaatiota voivat kuitenkin parittelun ja rekombinaation kautta päästä samanaikaisesti saman yksilön genomiin ja levitä samanaikaisesti populaatiossa. Edellä mainituista syistä seksuaalinen lisääntyminen nopeuttaa evoluutioprosessia. Maapallon biologisen evoluution yli kolmen miljardin vuoden aikana elämän kaksi ensimmäistä miljardia vuotta ovat pysyneet suvuttomassa lisääntymisvaiheessa, ja evoluutio on ollut hidasta, ja evoluutionopeus on kiihtynyt merkittävästi viimeisen miljardin vuoden aikana. Maapallon ympäristössä tapahtuneiden muutosten (kuten happea sisältävän ilmakehän syntyminen jne.) lisäksi myös sukupuolisen lisääntymisen esiintyminen ja kehittyminen on merkittävä syy.
Gametes
Gametes tarkoittaa sukusoluiksi kutsuttuja kypsiä soluja, joita syntyy lisääntymiselimistön tuottamana elimistön ollessa sukupuolisesti lisääntymässä. Sukusolut jaetaan uros-sukusoluihin ja naaras-sukusoluihin. Eläinten ja kasvien naaraspuolisia sukusoluja kutsutaan yleensä munasoluiksi ja urospuolisia sukusoluja siittiöiksi. Siittiöt ovat melko pieniä, mutta ne voivat liikkua ja tunkeutua munasoluun skorpionin muodossa. Munasolu on melko suuri ja uimataidoton. Esimerkiksi merisiilin munasolun tilavuus on 10 000 kertaa suurempi kuin siittiöiden. Vaikka uroksen ja naaraan sukusolut ovat tilavuudeltaan erilaisia, niiden jälkeläisille tarjoama ydin-DNA on yhtä suuri, eli kumpikin tarjoaa joukon genomeja. Munasolujen suuren koon vuoksi tytärsolujen sytoplasman rakenteen ja sytoplasman DNA:n tarjoavat kuitenkin pääasiassa munasolut. Sukusoluilla on tärkeä rooli biologisissa laskelmissa. Geenikarttojen avulla geenien kulkua ja jälkeläisten genotyyppien mallia voidaan tarkkailla selvästi. Sukusolujen lukumäärä hybridien ensimmäisessä sukupolvessa ja hybridien suhteelliset ominaisuudet ovat 2n:nnen potenssi (n on suhteellisten ominaisuuksien logaritmi). Kun elävä elimistö muodostaa sukusolun, parittaiset perintötekijät erotetaan toisistaan ja ne siirtyvät eri sukusoluihin. Sukusolut sisältävät kutakin perintötekijäparia vain yhden. Hedelmöitymisen yhteydessä uros- ja naaras-sukusolujen yhdistelmä on satunnainen. Meioosissa muodostuvien sukusolujen suvun, kromosomikoostumuksen moninaisuuden, eri sukusolujen perintöaineksen erilaisuuden sekä hedelmöittymisprosessissa tapahtuvan munasolujen ja siittiöiden yhdistymisen sattumanvaraisuuden vuoksi saman vanhemman jälkeläisten on oltava moninaisia.
Fertilisaatio
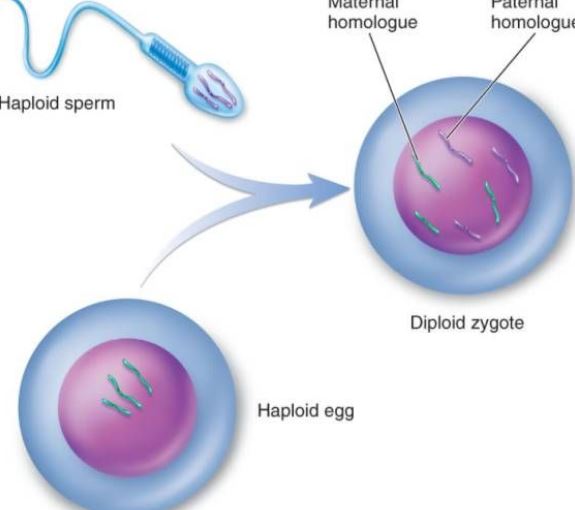
Fertilisaatio on prosessi, jossa munasolut ja siittiöt sulautuvat yhteen yhdeksi zygootiksi. Se on sukupuolisen lisääntymisen perusominaisuus, ja se on kaikkialla eläin- ja kasvimaailmassa, mutta siihen viitataan usein yleisimmin eläimiin. Eläinten lannoitus solutasolla lannoitusprosessi sisältää kolme päävaihetta: munasolun aktivoituminen, säätely ja amfoteerinen pronukleaarinen fuusio. Aktivointia voidaan pitää ontogeneesin alkupisteenä, joka ilmenee pääasiassa muutoksina tihkukalvon läpäisevyydessä, kortikaalisten rakeiden ekklesiassa, hedelmöittyneen kalvon muodostumisessa jne. Säätyminen tapahtuu aktivaation jälkeen, mikä on ensimmäinen vaihe hedelmöittyneiden munasolujen normaalin jakautumisen varmistamisessa. Prokaryoottisen ytimen muutos varmistaa vanhempien periytymisen ja palauttaa diploidin. Lannoitus ei ainoastaan käynnistä DNA:n monistumista, vaan myös aktivoi geneettisen informaation, kuten mRNA:n ja rRNA:n, munasolussa syntetisoimaan alkion kehityksessä tarvittavaa proteiinia. 6-7 päivän kuluttua hedelmöityksestä blastokystan zona pellucida katoaa ja vähitellen hautautuu ja peittyy kohdun limakalvoon, mitä kutsutaan hedelmöittyneen munasolun implantoitumiseksi. Eläimen siemennesteellä ei ole selvää kemotaksista kuten alempien kasvien, kuten bryofyyttien, siemennesteellä, vaan se pääsee munasolun läheisyyteen aktiivisella liikkeellä tai genitaaliepiteelisolujen sädekehän liikkeen avulla. Sperman kapasitaatio: Tiedetään, että kun monet nisäkkäiden siittiöt kulkevat naaraan lisääntymisväylän tai kumuluksen läpi, siittiöitä kapseloiva vieras proteiini poistuu ja siittiöiden plasmakalvon fysikaaliset ja biologiset ominaisuudet muuttuvat, jolloin siittiöt voivat saada energiaa ja osallistua hedelmöittymisprosessiin. Kun nisäkkäiden kapasitoitu siittiö koskettaa munasolukalvoa tai munasolun zona pellucidaa, se sitoutuu spesifisesti munasolukalvolla olevaan glykoproteiiniin, joka stimuloi siittiöitä tuottamaan energiaa. Akrosomireaktio auttaa siittiöitä ylittämään munasolukalvon edelleen. Merisiilin munasolussa siittiöiden akrosomireaktio on tietty polysakkaridiaine perikalvokalvolla. Useimmilla munilla on munakalvoja periferiassa, ja eri munakalvojen paksuus vaihtelee. Pääkomponentti on mukiini tai mukopolysakkaridi; vain harvat ovat paljaita munia, kuten koteloeläimen munat. Akrosomireaktio: Kun siittiö läpäisee munakalvon, tapahtuu sitoutumisprosessi. Ensin mainittu on löyhästi kiinnittynyt, vailla ulkoisia lämpötilahäiriöitä, eikä sillä ole spesifisyyttä. Tartunta-aikana akrosomaalisen kalvon protoplastiproteiini muuttuu akrosomiproteiiniksi, ja akrosomiproteiini kiihdyttää siittiöitä ylittämään munakalvon; Se on vahva yhdistelmä, matala lämpötila voi häiritä sitä, ja sillä on spesifisyys. Merisiilin siittiöiden plasmakalvolta on eristetty proteiini, joka sitoutuu spesifisesti munan glykoproteiiniin, jota kutsutaan sitovaksi proteiiniksi ja jonka molekyylipaino on noin 30 000. Ovulaation aikana toisen kypsymisjaon puolivälissä olevat sekundaariset munasolut yhdessä ympäröivän zona pellucidan ja sädekruunun kanssa kulkeutuvat nopeasti munanjohtimen ampulliin vatsaontelon kautta munanjohtimen epiteelisolujen värekarvojen heilahduksen ja lihaskerroksen supistumisen vuoksi. Jos munasolu ei kohtaa siittiöitä, se alkaa yleensä rappeutua 12-24 tunnin kuluessa. Kun siittiö on kosketuksissa munasolun kanssa, munasolu itse käy läpi joukon aktivoitumismuutoksia. Nisäkkäiden munasoluilla sille on ominaista kortikaalinen reaktio, munasarjakalvoreaktio ja zona pellucida -reaktio, jotka voivat estää monipuolista hedelmöittymistä ja stimuloida munasolujen jatkokehitystä. Kortikaalinen reaktio tapahtuu siittiöiden yhdistyessä. Fuusiokohdasta kuorihiukkaset repeävät ja sisältö purkautuu, jolloin koko munasolun kuori leviää. Munasolukalvoreaktio on munasolun ja kortikaalisen rakeen kuoren rekombinaatioprosessi. Zona pellucida -reaktio on prosessi, jossa kortikaalinen effluksi ja zona pellucida irtoavat hedelmöittyneestä kalvosta, munasolukalvo irtoaa plasmakalvosta, siittiöiden reseptori zona pellucidassa katoaa ja zona pellucida kovettuu. Vain siittiöt, joilla on akrosomireaktio, voivat sulautua munasolun kanssa. Akrosiinin vaikutuksesta siittiö kulkee sädekruunun läpi ja vuorovaikuttaa zona pellucidassa olevan siittiöreseptorin glykoproteiinimolekyylin ZP3:n kanssa, jolloin siittiö vapauttaa akrosomaalisen entsyymin ja pääsee perivitelliniin zona pellucidan kautta. Hedelmöityksen alussa ihmisen siittiöiden pään ekvatoriaalisen osan kuori on kosketuksissa munasolukalvon kanssa, minkä jälkeen siittiöiden tuma ja sytoplasma pääsevät munasoluun. Siittiöiden tultua munasoluun munasolun matalassa sytoplasmassa olevat kuorihiukkaset vapauttavat välittömästi sisältönsä kalvon periferiaan. Samalla, kun kortikaalirakeiden kalvo sulautuu munasolujen kanssa, solun pinnan negatiivinen pintavaraus kasvaa, mikä estää siittiöiden plasmakalvon sulautumisen munasolukalvoon, mitä kutsutaan kortikaalireaktioksi. Zona pellucida -rakenteen muutoksia kutsutaan zona pellucida -reaktioksi. Tällöin zona pellucidan kyky sitoa siittiöitä vähenee, mikä estää polyspermian syntymisen ja varmistaa ihmisen monospermian biologiset ominaisuudet. Sen lisäksi, että kortikaalisten rakeiden sisältö tuhoaa tai inaktivoi siittiöihin sitoutuneen reseptorin zona pellucidassa, se voi myös lisätä peptidiketjujen ristisilloittumista zona pellucidassa, heikentää herkkyyttä akrosomiproteaasille ja estää siittiöiden tunkeutumisen. Vaikka useita siittiöitä kulkee zona pellucidan läpi, vain yksi siittiö tunkeutuu munasoluun hedelmöittämään sitä. Epänormaalissa tilanteessa hedelmöitykseen osallistuu kaksi siittiöitä, eli kyseessä on kaksinkertainen siittiöiden hedelmöittyminen. Kaksi siittiöitä tunkeutuu samanaikaisesti munasoluun muodostaen triploidien solujen alkioita, jotka kaikki abortoituvat tai kuolevat pian syntymän jälkeen. Siittiöiden päästyä munasoluun munasolu suorittaa nopeasti toisen kypsän jakautumisen. Tällöin siittiöiden ja munasolun ytimiä kutsutaan uroksen pronukleukseksi ja naaraan pronukleukseksi. Molemmat pronominit lähestyvät vähitellen toisiaan, ydinkalvo katoaa ja kromosomit sulautuvat yhteen muodostaen diploidisen hedelmöittyneen munasolun. Kun siittiösolut ovat fuusioituneet, munasolun pinnalla olevien mikrovillien voidaan nähdä ympäröivän siittiöitä, jotka voivat olla suuntautuneita; sitten munasolukalvo fuusioituu plasmakalvon kanssa siittiöiden akrosomin takimmaisella alueella.
Viittaus
- Yanagimachi R. Sukusolut ja hedelmöittyminen: miksi tutkin näitä aiheita, ja mitä opin tutkimukseni varrella. Andrologia. 2014, 2(6):787-93.
- Henshaw J M, Marshall D J, Jennions M D, et al. Local gamete competition explains sex allocation and fertilization strategies in the sea. American Naturalist. 2014, 184(2):32.
- Mori T, Igawa T. Gamete attachment process revealed in flowering plant fertilization. Plant Signaling & Behavior. 2014, 9(12):977715.
- Hédouin L, Pilon R, Puisay A. Hyposalinity stressi vaarantaa enemmän sukusolujen hedelmöittymistä kuin korallitoukkien selviytymistä. Marine Environmental Research. 2015, 104:1-9.
Luo J, Mcginnis L K, Carlton C, et al. PTK2b:n toiminta hiiren munasolun hedelmöityksen aikana. Biochem Biophys Res Commun. 2014, 450(3):1212-1217.