- Volvox Case Study Guide Covers up Classification, Characteristics, Volvox Under Microscope, Reproduction (Asexual and Sexual Reproduction)
- Tarkka opas kaikkine näkökohtineen
- Volvokit Luokitus
- Volvoxin ominaisuudet
- Kasvurunko (Gametofyytti)
- Solujen rakenne
- Volvox yksisoluinen tai monisoluinen
- Volvox mikroskoopilla
- Volvoxin lisääntyminen
- Sukupuolinen lisääntyminen
- Sukupuolista lisääntymistä
Volvox Case Study Guide Covers up Classification, Characteristics, Volvox Under Microscope, Reproduction (Asexual and Sexual Reproduction)
Volvox on makean veden planktoninen (vapaasti kelluva) levä. Näihin sukuihin kuuluu noin 20 lajia. Kasvien valtakunnassa se esiintyy kauneimpana ja houkuttelevimpana esineenä. Sitä esiintyy makeassa vedessä nuppineulanpään kokoisina vihreinä palloina. Ne ovat juuri ja juuri paljain silmin havaittavissa, halkaisijaltaan noin 0,5 mm.
Tarkka opas kaikkine näkökohtineen
Erityisen uintikuvionsa vuoksi levästä käytetään usein nimitystä rullaava levä. Sen kasvua havaitaan usein tilapäisissä tai pysyvissä makean veden lammikoissa, altaissa, ojissa ja myös järvissä. Kasvu on runsasta, kun lämpötilaa ja orgaanista ainesta on riittävästi saatavilla. Sen lisääntyminen on niin yleistä ja runsasta, että lampien vesi muuttuu vihreäksi (vesikukka). Kevät- ja sadekaudet ovat sen aktiivisen kasvullisen kasvun tavanomaisia kausia. Epäsuotuisan jakson (kesä) alkaessa levä häviää ja kulkee epäsuotuisan jakson zygootin muodossa. Volvokit valmistavat oman ravintonsa fotosynteesin avulla.
Volvokit Luokitus
- Luokka: Luokka: Chlorophyceae
- Järjestys: Volvocales
- Alalahko: Chlamydomonadineae
- Suku: Volvox
Yleisimmät Volvox-lajit ovat V, globator, V, aureus; V. prolificus, V. rouseletti, V. merelli.
Volvoxin ominaisuudet
Kasvurunko (Gametofyytti)
Volvox on koenobiaalinen viherlevä, {(kolonia-kasvurunko ei ole kiinteä solujen lukumäärä esim. Volvox aureus) (koenobium-kasvurunko, jolla on kiinteä solujen lukumäärä, esim, Pandorina moruma, solujen lukumäärä on 4, 8, 16 tai 32. Eudorina unicocca, solujen lukumäärä 16, 32 tai 64)}. Liikkuvista muodoista Volvoxin coenobium-levät ovat suurimpia, pitkälle erilaistuneita ja hyvin kehittyneitä leviä. Kukin coenobium on ellipsinmuotoinen tai ontto pallomainen runko, jossa on tarkasti merkitty herkkä limakalvokerros. Koenobiumin sisäosa koostuu diffluenttisesta (vesipitoisesta) limaaineesta, kun taas solut ovat järjestäytyneet yhdeksi kerrokseksi periferiassa.
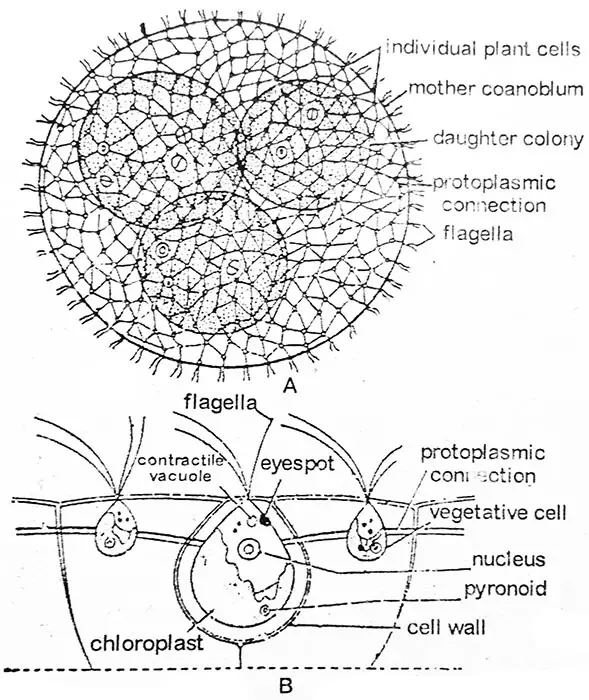
Pesäkkeen liikkeen saa aikaan yksittäisten solujen lippulaivojen yhteinen toiminta. Jokaisella coenobiumilla on selvä etu- ja takapää. Koenobiumissa on polariteetti, se liikkuu ja pyörii hitaasti ja osoittaa huomattavaa yhteistyötä etu- ja takapään solujen välillä liikkeensä aikana. Volvox ei ole yksilö vaan useiden samankaltaisten solujen yhteenliittymä, joista kukin toimii kuin itsenäinen yksilö ja huolehtii omasta ravinnostaan, hengityksestään ja erittymisestään, eikä solujen välillä ole yhteistyötä näissä toiminnoissa. Solujen lukumäärä koenobiaa kohti vaihtelee esim. 500-1000 V. aureuksessa, 1500-20 000 V. globatorissa ja jopa noin 60 000:een V. rouselettissä.
Solujen rakenne
Tutkimusten mukaan jokaisella yksittäisellä solulla on oma identiteettinsä. Sitä ympäröi oma suuri hyytelömäinen, hyytelömäinen vaippansa, joka voi olla ristiriidassa viereisten solujen vaippojen kanssa tai se voi olla toisistaan erillinen. Silloin ne ovat erillisiä, ne ovat kulmikkaita keskinäisen puristumisen vuoksi ja ovat yleensä ääriviivoiltaan kuusikulmaisia. Kuva,2.22. Näin ollen huomattava määrä hyytelömäistä materiaalia auttaa erottamaan yhden solun toisesta solusta. Useimmissa lajeissa kukin solu on yhteydessä naapurisoluihinsa protoplasma- tai sytoplasmasäikeillä (plasmodesmae, kuva 2.22), jotka ovat syntyneet solun jakautumisen ja pesäkkeen kehittymisen aikana. Protoplasmasäikeet voivat olla ohuita ja hentoja V. aureuksessa, pyöreitä V. globatorissa tai kiilamaisia V. mononaessa, tai ne voivat puuttua, kuten V. tertiuksessa.
Volvoxin yksittäinen solu muistuttaa pääpiirteissään Chlamydomopasia. Kussakin solussa on etupuolella parin verran yhtä pitkiä lippulankoja. Molemmat lippulat ovat piiskatyyppisiä. Lippulangat työntyvät coenobiumin pinnan ulkopuolelle ympäröivään veteen. Lippuloiden tyven lähellä on kaksi tai useampia supistuvia vakuoleja. Protoplasti on muodoltaan erilainen lajista riippuen. V. tertius -bakteerin protoplasti on pyöreä ja Chlamydomonas-tyyppinen, kun taas V. globator -bakteerin protoplasti on tähtimäinen, ja siinä on diffuusia kloroplastia ja hajallaan olevia supistuvia vakuoleja.
Volvox yksisoluinen tai monisoluinen
Nuoressa pesäkkeessä vegetatiiviset solut ovat vihreitä ja kooltaan ja muodoltaan samankaltaisia, mutta vanhemmissa pesäkkeissä tietyt takimmaisen alueen solut kasvavat kymmenkertaisiksi tai suuremmiksi kuin normaali pesäkkeen solun koko. Ne kehittävät lukuisia pyrenoideja, joiden koko kasvaa ja jotka toimivat lisääntymissoluina. Ne voivat olla suvuttomia tai seksuaalisia. Joissakin tapauksissa samassa pesäkkeessä voi olla sekä suvuttomia että sukusoluja. Pesäkkeen etuosassa solut ovat vain kasvullisia ja verrattain pienempiä. Etuosan soluissa on suurempi silmäpiste. Pesäkkeessä on siis kahdenlaisia soluja: lisääntymissoluja ja somaattisia soluja.
Volvox voi toimia esimerkkinä ensimmäisestä askeleesta kohti koordinointia ja työnjakoa. Volvoxin pesäkettä voidaan pitää monisoluisena tyyppinä, joka koostuu eri toimintojen suorittamista varten erotetuista soluista. Eri tehtäviä suorittavia soluja ovat,
- Vegetatiiviset solut, jotka liittyvät ravinnon valmistukseen ja osallistuvat liikkumiseen,
- Sukupuoliset solut, jotka tuottavat tytärpesäkkeitä
- Sukupuoliset solut: tuottavat munia ja tuottavat antheridiumia
Volvox mikroskoopilla
Alhaalla on mikroskooppikuva volvoxin pesäkkeestä:
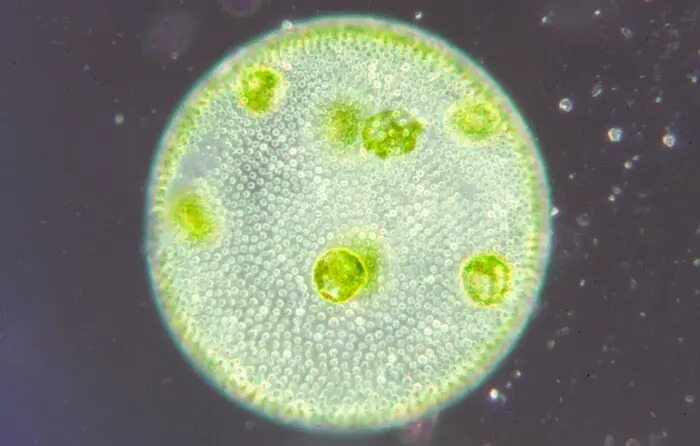
Volvoxin lisääntyminen
Vastoin kuin Chlamydomonasissa volvoxin pesäkkeen soluissa on havaittavissa funktionaalista erikoistumista. Se lisääntyy sekä suvuttomasti että sukupuolisesti. Kasvukauden alussa (suotuisat olosuhteet). lisääntyminen on suvutonta. Tästä syystä kaikki kerrallaan kerätyt pesäkkeet ovat joko suvuttomia tai sukupuolisia.
Sukupuolinen lisääntyminen
Sukupuolinen lisääntyminen tapahtuu kasvukauden alussa suotuisissa olosuhteissa. Aikaisemmissa vaiheissa kaikki pesäkkeen solut ovat samanlaisia, mutta myöhemmin muutamat pesäkkeen takimmaisessa puoliskossa olevat solut varastoivat ravintoa ja kasvavat kooltaan. Nämä huomattavasti suurentuneet solut ovat erikoistuneita suvuttomia soluja, joita kutsutaan gonidioiksi (yksikkö gonidium). Kukin gonidio sijaitsee pallomaisen hyytelömäisen tupen sisällä.
Gonidiaalisen protoplastin ensimmäinen jakautuminen on pitkittäinen eli coenobiumin anterio-posteriorinen taso. Toinen jakautuma on myös pitkittäinen ja suorassa kulmassa ensimmäiseen nähden. Kukin näin muodostuneista tytärsoluista jakautuu jälleen pituussuunnassa niin, että muodostuu kahdeksan solun levy. Se kaartuu siten, että sen kovera pinta on ulospäin. Tätä kahdeksan solun vaihetta kutsutaan Plakea-vaiheeksi. Tytärsolujen samanaikaiset pituussuuntaiset jakautumiset jatkuvat useiden solusukupolvien ajan (V. rouselettillä jopa 14, 15 tai 16 kertaa).
Kuudentoista solun vaiheessa solut ovat järjestäytyneet onton pallon kehälle, jossa on pieni aukko, phialopore, vanhemman coenobiumin ulkopuolelle päin. Tässä vaiheessa kaikki solut ovat paljaita ja kosketuksissa toisiinsa. Niiden etupäät osoittavat kohti pallon keskustaa. Invaginaation edetessä fialopori laajenee huomattavasti. Kun takimmaisen osan sisäänpäin taittuminen (invaginaatio) alkaa työntyä phialoporeen, sen ympäröivät reunat käpristyvät taaksepäin ja liukuvat vähitellen alaspäin, kunnes koko rakenne on kääntynyt ylösalaisin. Fialopori, jossa on nyt useita poimuja, sulkeutuu vähitellen. Inversioprosessi kestää noin kolmesta viiteen tuntia. Joissakin epänormaaleissa tapauksissa inversiota ei tapahdu lainkaan, kuten on raportoitu V. minorissa.
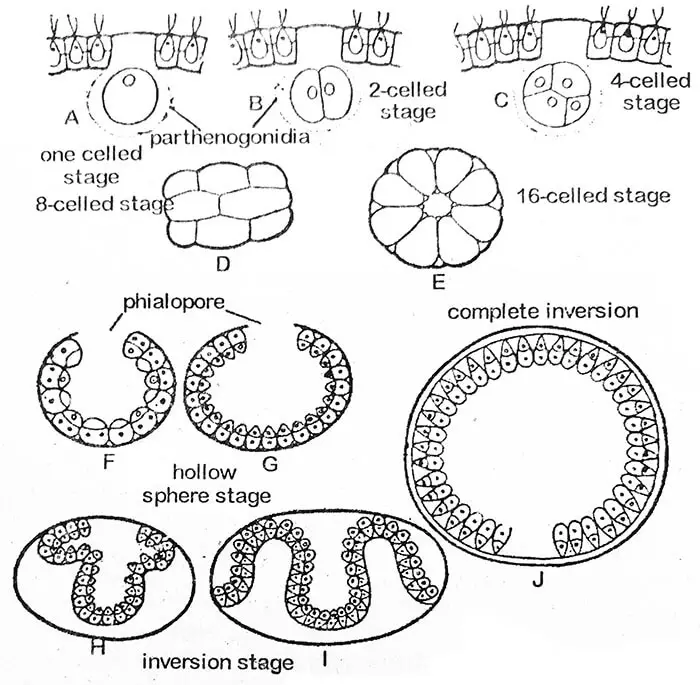
Tytärpesäkkeiden solut alkavat nyt erottua toisistaan kehittämällä limaisia osia (soluseinää). Kukin solu saa lopulta lippalakiparin ja solukalvon. Tytärpesäke (coenobium) säilyy edelleen vanhemman soluseinän sisällä, joka lopulta kehittyy sitä ympäröiväksi limakalvoksi. Vanhempaan pesäkkeeseen voi kehittyä samanaikaisesti useita tytärkoenobioita. Näin ne voivat täyttää emopesäkkeen onton keskialueen.
Tytärcoenobiat vapautuvat emopesäkkeen kuollessa ja hajotessa. Joskus tytärpesäkkeet eivät vapaudu pidemmäksi aikaa ja kehittävät tytärpesäkkeitä. Siten yhdessä alkuperäisessä emopesäkkeessä voi esiintyä jopa 2-4 sukupolvea vangittuja tytärpesäkkeitä, erityisesti V. africanuksella.
Sukupuolista lisääntymistä
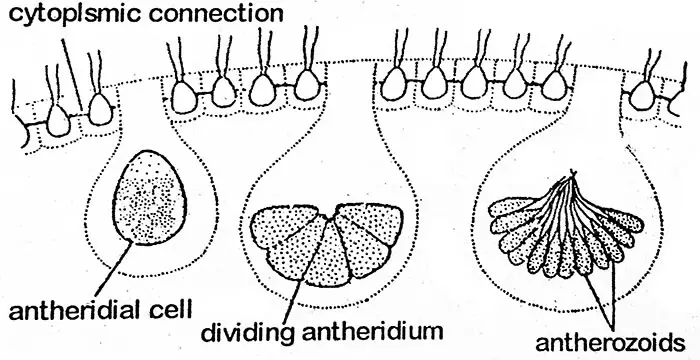
Volvoxilla on kehittynyt oogamotyyppinen sukupuolinen lisääntyminen, joka tapahtuu antheridioiden ja oogonioiden muodostumisen kautta. Ne voivat muodostua samaan coenobiumiin (yksikotinen) kuten V. globatorilla tai eri coenobiumiin (kaksikotinen) kuten V. aureuksella. Yksikotiset lajit ovat protandroosia (antheridia kehittyy ensin), joten tällaisissa lajeissa hedelmöittyminen tapahtuu eri kasvien antherozoidin ja munasolun välillä. On varsin mielenkiintoista, että suvulliset pesäkkeet ovat usein vailla suvuttomasti muodostuneita tytärpesäkkeitä. Koenobiumissa sukupuolielimen muodostamiseen tarkoitetut solut ovat takimmaisessa puoliskossa. Niitä pidetään erikoistuneina soluina. Sukuelimiä (gametangioita) syntyy vähemmän. Gametangioiden muodostumisen aikana solu suurenee ja pyöristyy ja hylkää lippulaput, mutta se pysyy yhteydessä muihin soluihin hienoilla protoplasmisilla säikeillä. Uroksen gametangiaa kutsutaan antheridiumiksi, kun taas naaraan oogoniumiksi.
Antheridiumin protoplasti käy läpi toistuvia solunjakautumia samalla tavalla kuin havaitaan suvuttoman gonidiaalisolun kehittyessä tytärpesäkkeeksi (eli plakea-vaiheessa). Näin muodostuu massa. pieniä, alastomia, biflagellaattisia, fusiformisia antherozoideja, joiden lukumäärä antheridiumissa on 16-512. Nämä ryhmittyvät litteäksi levyksi lukuun ottamatta V. aureusta, jossa anterotsoidit näkyvät suvuttomassa pesäkkeessä. Antherozoidien plakea dissosioituu ja vapauttaa antherozoidit.
Antherozoidi
Jokainen antherozoidi on kaksisoluinen, pitkänomainen, kartiomainen tai fusiforminen rakenne, jossa on yksittäinen ydin ja pieni kellanvihreä tai vaaleanvihreä kloroplasti.
Oogonium
Oogoniaalisolu suurenee huomattavasti ja hylkää lippulankansa ja protoplasmaattiset yhteytensä naapurisoluihin. Solu muuttuu pyöreäksi tai pullonmuotoiseksi, ja suuri osa sen osasta työntyy coenobiumin sisälle. Tässä vaiheessa sitä kutsutaan oogoniumiksi, jonka koko osa muuttuu yhdeksi pallomaiseksi munaksi, jonka toisella puolella on nokkamainen uloke. Tämän pään kautta anterotsoidi tunkeutuu oogoniumiin. Munassa (oosfäärissä) on suuri, keskellä sijaitseva tuma ja parietaalinen kloroplasti, jossa on pyrenoideja. Siihen on varastoitunut runsaasti varoaineita, jotka usein imeytyvät naapurisoluista protoplasmasäikeiden kautta.
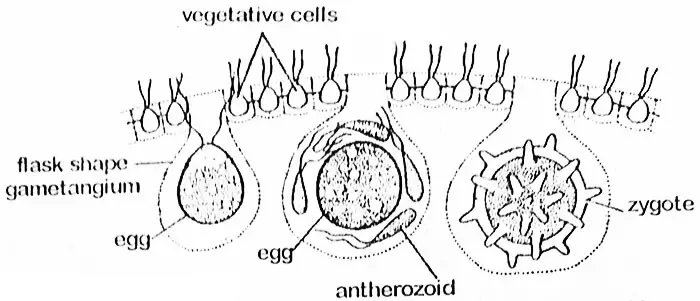
Fertilisaatio
Fertilisaation yhteydessä anterotsoidit vapautuvat ryhmittäin, ja nämä pysyvät koskemattomina munasolun saavuttamiseen asti. Sen jälkeen anterotsoidit vapautuvat. Vain yksi anterotsoidi sulautuu munasolun kanssa ja johtaa oospoorin muodostumiseen. Oospori erittää myöhemmin kolmikerroksisen sileän tai piikkiseinämän. Se kerää riittävästi hematokromia (punaisia väripigmenttirakeita, jotka ovat todennäköisesti luonteeltaan ksantofylliä), mikä antaa sille oranssin värisen ulkonäön. Tässä vaiheessa sitä voidaan kutsua zygootiksi.
Oospori ja sen itäminen
Ulkoseinämä, eksospori, voi olla sileä (V. globator) tai piikkinen (V. speematospaera). Keskimmäinen kerros on mesospori ja sisempi on endospori. Zygootti sisältää riittävästi vararavintomateriaalia ja muita sulkeumia. Näin ollen zygootti säilyy coenobiumissa, joka voi vapautua hyytelömäisen matriisin hajotessa. Vapautumisen jälkeen se laskeutuu altaan pohjalle ja voi säilyä elinkelpoisena useita vuosia.

Suotuisten olosuhteiden ilmaantuessa zygootti kehittyy eri tavoin. V. campensiksessa zygootin ydin jakautuu meioottisesti ja muodostaa neljä ydintä, joista kolme degeneroituu ja yksi jää eloon: Eloonjäänyt ydin ja sytoplasman sisältö poistuvat vesikkelistä. Tässä vaiheessa sitä voidaan kutsua swarmeriksi (suuri määrä tai tiheä ryhmä hyönteisiä, soluja jne.), se ui vapaasti ja jakautuu ja jakautuu uudelleen muodostaakseen uuden coenobiumin. Itämisen aikana kaksi ulompaa seinämäkerrosta muuttuvat hyytelömäisiksi ja sisempi kerros muodostaa vesikkelin, joka myöhemmin täyttyy zygoottiprotoplastilla. V. rouselettissä ja V. minorissa zygootin protoplasti muuttuu yhdeksi zoosporaksi, joka edelleen jakautumalla muodostaa uuden coenobiumin. Tällainen coenobium koostuu pienemmästä määrästä soluja, jotka lisääntyvät suvuttomasti seuraavien kuuden tai useamman sukupolven ajan, ja niiden määrä kasvaa joka kerta seuraavissa sukupolvissa.
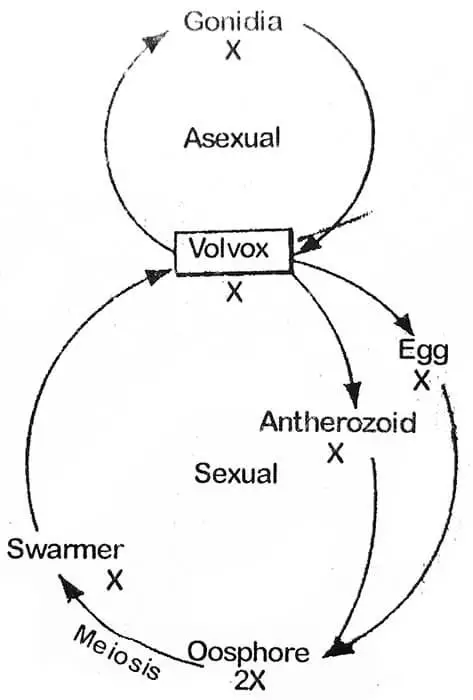
Zygootti on ainoa diploidi vaihe Volvoxin elinkierrossa, ja näin ollen kasvin päärunko on haploidi. Siksi zygootti joutuu käymään läpi pelkistävän jakautumisen uutta pesäkettä muodostaessaan.
Hyvinvointikykyinen kasvi.