Gameták és megtermékenyítés áttekintése
A szexuális reprodukció a szülők által termelt ivarsejtek (ivarsejtek), a kétnemű ivarsejtek (például a spermium és a petesejt) kombinációja révén a megtermékenyített petesejt lesz, majd a megtermékenyített petesejtből egy új egyed fejlődik, amit szexualitás reprodukciónak nevezünk. A szexuális szaporodás során a génkombinációk kiterjedt variációja növelheti az utódok képességét a természetes szelekcióhoz való alkalmazkodásra. Az ivaros szaporodás utódaiban véletlenszerűen kombinált gének vagy előnyösek a faj számára, vagy nem, de legalábbis növelik annak esélyét, hogy néhány egyed túléljen a kiszámíthatatlan és állandóan változó környezetben, és ezáltal a faj javát szolgálják. A szexuális szaporodás elősegítheti a kedvező mutációk terjedését is a populációban. Ha egy faj két egyedének különböző helyeken vannak kedvező mutációi, az aszexuális szaporodású populációban a két mutáns addig versenyez egymással, amíg az egyik ki nem szűnik, lehetetlen mindkét kedvező mutációt egyszerre megtartani. A szexuális szaporodású populációkban azonban a párosodás és a rekombináció révén ez a két kedvező mutáció egyszerre kerülhet ugyanannak az egyednek a genomjába, és egyszerre terjedhet el a populációban. A fenti okok miatt a szexuális szaporodás felgyorsítja az evolúció folyamatát. A földi biológiai evolúció több mint 3 milliárd éve alatt az élet első kétmilliárd éve az aszexuális reprodukciós szakaszban maradt, és az evolúció lassú volt, az utolsó 1 milliárd év óta pedig jelentősen felgyorsult az evolúció üteme. A földi környezet változásai mellett (például az oxigéntartalmú légkör kialakulása stb.) az ivaros szaporodás megjelenése és fejlődése is fontos ok.
Gaméták
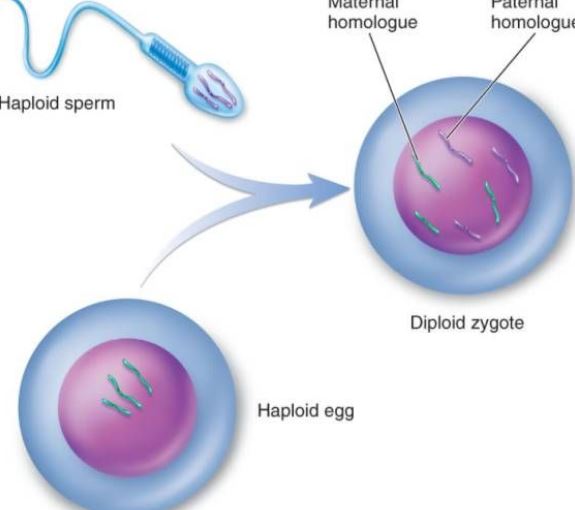
A gaméták a szervezet ivaros szaporodásakor a szaporodási rendszer által termelt érett sejteket jelentik, amelyeket csírasejteknek nevezünk. A gaméták hímivarsejtekre és nőivarsejtekre oszthatók. Az állatok és növények női ivarsejtjeit általában petesejteknek, a hímivarsejteket pedig spermiumoknak nevezik. A spermium meglehetősen kicsi, de képes mozogni, és skorpió alakban behatolni a petesejtbe. A petesejt meglehetősen nagy és nem úszik. Például a tengeri sün petesejtjének térfogata 10 000-szer nagyobb, mint a spermiumoké. Bár a hím és a nőstény ivarsejtek térfogata különbözik, az általuk az utódok számára biztosított nukleáris DNS azonos, azaz mindkettő egy-egy genomkészletet biztosít. A petesejtek nagy mérete miatt azonban a leánysejtek citoplazmatikus szerkezetét és citoplazmatikus DNS-ét alapvetően a petesejtek biztosítják. Az ivarsejtek fontos szerepet játszanak a biológiai számításokban. A genetikai térképeken keresztül a gének áramlása és az utódok genotípusainak mintázata egyértelműen megfigyelhető. A hibridek első generációjában az ivarsejtek száma és a hibridek relatív tulajdonságai 2n-edik hatványa (n a relatív tulajdonságok logaritmusa). Amikor egy élőlény ivarsejtet képez, a párosított genetikai tényezők elválnak egymástól, és különböző ivarsejtekbe kerülnek. Az ivarsejtek minden genetikai faktorpárból csak egyet tartalmaznak. Megtermékenyítéskor a hím és női ivarsejtek kombinációja véletlenszerű. A meiózis során kialakuló ivarsejtek nemzetségének, a kromoszómaösszetétel változatosságának, a különböző ivarsejtek genetikai anyagának különbözőségének, valamint a megtermékenyítés során a petesejtek és a spermiumok kombinációjának véletlenszerűségének köszönhetően az azonos szülőktől származó utódoknak változatosnak kell lenniük.
Fertilizáció
A megtermékenyítés az a folyamat, amelynek során a petesejt és a spermium egy zigótává olvad össze. Ez az ivaros szaporodás alapvető jellemzője, és mindenütt jelen van az állat- és növényvilágban, de a leggyakrabban az állatvilágban hivatkoznak rá. Az állati megtermékenyítés sejtszinten a megtermékenyítési folyamat három fő szakaszból áll: a petesejt aktiválása, a szabályozás és az amfoterikus pronukleáris fúzió. Az aktiválást tekinthetjük az ontogenezis kiindulópontjának, amely főként a petesejtmembrán áteresztőképességének változásaiban, a kéregszemcsék ekkléziájában, a megtermékenyített membrán kialakulásában stb. nyilvánul meg. Az alkalmazkodás az aktiválódás után következik be, ami az első lépés a megtermékenyített petesejtek normális osztódásának biztosításában. A prokarióta sejtmag megváltozása biztosítja a szülők öröklődését és visszaállítja a diploidot. A megtermékenyítés nemcsak a DNS replikációját indítja el, hanem a genetikai információt, például az mRNS-t és az rRNS-t is aktiválja a petesejtben az embrió fejlődéséhez szükséges fehérje szintéziséhez. A megtermékenyítést követő 6-7 nap elteltével a blasztociszta zona pellucida eltűnik, és fokozatosan betemetődik, és a méhnyálkahártya borítja, amit a megtermékenyített petesejt beágyazódásának nevezünk. Az állati spermiumok nem rendelkeznek nyilvánvaló kemotaxissal, mint az alacsonyabb rendű növények, például a bryophyták spermiumai, hanem inkább aktív mozgással vagy a genitális hámsejtek csillószőrös mozgása révén jutnak el a petesejt közelébe. Spermakapacitáció: Ismert, hogy amikor sok emlős spermium áthalad a női reproduktív traktuson vagy a cumuluson, a spermiumot beburkoló idegen fehérje eltávolodik, és a spermium plazmamembránjának fizikai és biológiai tulajdonságai megváltoznak, lehetővé téve a spermium számára, hogy energiát nyerjen és részt vegyen a megtermékenyítési folyamatban. Amikor az emlősök kapacitált spermiuma érintkezik a petesejt membránjával vagy a petesejt zona pellucidájával, specifikusan kötődik a petesejt membránján lévő glikoproteinhez, ami a spermiumot energiatermelésre serkenti. Az akroszómareakció segíti a spermiumot abban, hogy tovább haladjon a petesejt membránján. A tengeri süntojáson a spermiumok akroszómareakcióját egy bizonyos poliszacharid anyag a perimembrán filmben. A legtöbb petesejtnek a perifériáján van petesejtmembránja, és a különböző petesejtmembránok vastagsága változó. A fő összetevő a mucin vagy a mukopoliszacharid; csak néhány tojás csupasz, mint például a bélpoklosok petéi. Akroszómareakció: Amikor a spermium áthalad a petesejt membránján, kötési folyamat megy végbe. Az előbbi laza kötődésű, külső hőmérsékleti zavaroktól mentes, és nem specifikus. A tapadási időszak alatt az akroszóma membránon lévő protoplaszt fehérje akroszóma fehérjévé alakul át, és az akroszóma fehérje gyorsítja a spermiumot a petesejt membránon való áthaladásban; Erős kötés, alacsony hőmérséklet zavarhatja, és specifitással rendelkezik. A tengeri sün spermiumok plazmamembránján izoláltak egy olyan fehérjét, amely specifikusan kötődik a petesejt glikoproteinjéhez, az úgynevezett kötőfehérjét, amelynek molekulatömege kb. 30 000. Az ovuláció során a második érési osztódás közepén lévő másodlagos petesejtek a környező zona pellucidával és a sugárkoronával együtt a petevezető hámsejtek csillóinak kilengése és az izomréteg összehúzódása következtében a hasüregen keresztül gyorsan bejutnak a petevezető ampullába. Ha a petesejt nem találkozik a spermiummal, általában 12-24 órán belül degenerálódni kezd. Amint a spermium kapcsolatba kerül a petesejttel, maga a petesejt egy sor aktivációs változáson megy keresztül. Az emlősök petesejtjénél ez a kéregreakció, a petefészekhártya-reakció és a zona pellucida-reakció jellemzi, amelyek megakadályozhatják a többszörös megtermékenyülést és serkenthetik a petesejt további fejlődését. A kérgi reakció a spermiumok összeolvadásakor következik be. A fúzió pontjától a kéregrészecskék felrepednek és a tartalmuk kiáramlik, ezáltal a teljes petesejt kéregállománya szétterül. A petefészekhártya-reakció a petesejt és a kéregszemcsehalmaz rekombinációs folyamata. A zona pellucida reakció egy olyan folyamat, amelyben a megtermékenyített membránból a kérgi kiáramlás és a zona pellucida, a petesejt membránja elválik a plazmamembrántól, a zona pellucidában lévő spermiumreceptor eltűnik, és a zona pellucida megkeményedik. Csak az akroszómareakciót végző spermiumok tudnak összeolvadni a petesejttel. Az akrozin hatására a spermium áthalad a radiális koronán, és kölcsönhatásba lép a zona pellucidán lévő ZP3 spermareceptor glikoprotein molekulával, így a spermium felszabadítja az akroszómális enzimet, és a zona pellucidán keresztül belép a perivitellinbe. A megtermékenyítés kezdetén az emberi spermiumfej ekvatoriális részének burka érintkezik a petesejt membránjával, majd a spermium sejtmagja és citoplazmája belép a petesejtbe. Miután a spermium belép a petesejtbe, a petesejt sekély citoplazmájában lévő kéregrészecskék azonnal kibocsátják tartalmát a membrán perifériájára. Ugyanakkor, ahogy a kéregszemcsék membránja összeolvad a petesejtekkel, a sejtfelszínen megnő a negatív felületi töltés, ezáltal elnyomja a spermium plazmamembránjának a petesejt membránjával való összeolvadását, amit kéregreakciónak nevezünk. A zona pellucida szerkezetének megváltozását zona pellucida-reakciónak nevezzük. Ekkor a zona pellucida spermiummegkötő képessége csökken, megakadályozva a poliszpermia kialakulását és biztosítva az emberi monospermia biológiai jellemzőit. A zona pellucidában a spermiumokhoz kötött receptor elpusztítása vagy inaktiválása mellett a kéregszemcsék tartalma növelheti a zona pellucida peptidláncai közötti keresztkötést, gyengítheti az akroszóma proteázzal szembeni érzékenységet, és megakadályozhatja a spermiumok behatolását. Bár több spermium is áthalad a zona pellucidán, csak egy spermium jut be a petesejtbe, hogy megtermékenyítse azt. Abnormális esetben két spermium vesz részt a megtermékenyítésben, azaz kettős spermium megtermékenyítésről van szó. Két spermium egyszerre lép be a petesejtbe, és triploid sejtekből álló embriókat képeznek, amelyek mindegyike elvetél vagy születés után hamarosan elpusztul. Miután a spermium belép a petesejtbe, a petesejt gyorsan befejezi a második érett osztódást. Ekkor a spermium és a petesejt magját hímivarsejtmagnak, illetve női magnak nevezik. A két pronukleusz fokozatosan közeledik egymáshoz, a maghártya eltűnik, és a kromoszómák összeolvadnak, így egy diploid megtermékenyített petesejt jön létre. Amikor a spermiumok összeolvadnak, a petesejt felszínén látható, hogy a mikrovillák körülveszik a spermiumot, amely orientálódhat; ezután a petesejt membránja összeolvad a plazmamembránnal a spermium akroszóma hátsó régiójában.
Hivatkozás
- Yanagimachi R. Germ cells and fertilization: Why I studied these topics and what I learned along the path of my study. Andrológia. 2014, 2(6):787-93.
- Henshaw J M, Marshall D J, Jennions M D, et al. Local gamete competition explains sex allocation and fertilization strategies in the sea. American Naturalist. 2014, 184(2):32.
- Mori T, Igawa T. Gamete attachment process revealed in flowering plant fertilization. Plant Signaling & Behavior. 2014, 9(12):977715.
- Hédouin L, Pilon R, Puisay A. Hyposalinity stressz jobban veszélyezteti az ivarsejtek megtermékenyülését, mint a koralllárvák túlélését. Marine Environmental Research. 2015, 104:1-9.
Luo J, Mcginnis L K, Carlton C, et al. PTK2b funkció az egér petesejt megtermékenyítése során. Biochem Biophys Res Commun. 2014, 450(3):1212-1217.