ADVERTISEMENTS:
この記事では、ゾウリムシについて説明します: – 1.生息地、生息地と文化 ゾウリムシの2.運動量. ゾウリムシの構造 3. 運動量 4. 栄養 5. 呼吸と排泄 6. 体温調節 7. 行動 8. 生殖 9. 生殖における異常行動 10. いくつかの細胞質粒子
内容物。
- 生態,
- ゾウリムシの構造と培養
- ゾウリムシの運動
- ゾウリムシの栄養
- 呼吸と排泄
- 体温調節
- ゾウリムシの行動
- ゾウリムシの繁殖
- ゾウリムシの繁殖における異常な行動
- 生息地:ゾウリムシの細胞質微粒子報告 Habit, Paramecium Caudatumの生息地と培養。
- ゾウリムシの培養。
- ゾウリムシの構造:
- (i) ゾウリムシの大きさと形:
- (ii)ペリクル。
- (iii) 繊毛。
- (iv)毛細血管内系。
- (v)口腔溝とサイトピージ。
- (vi) 細胞質:
- (vii) Ectoplasm:
- (viii) トリコシスト(Trichocysts)。
- (ix) 神経原線維と関連原線維(Paramecium and Associated Fibrils)
- (x) 内膜:
- (xi) 収縮性液胞:
- (xii)食液胞。
- ゾウリムシの運動:
- (i)メタボリックまたは体の歪み。
- (ii) Ciliary Locomotion(繊毛運動)。
- Paramecium Caudatumの栄養:
- 摂食のしくみ。
- Paramecium Caudatumの消化と排泄:
- ゾウリムシの呼吸と排泄:
- Paramecium Caudatumの行動:
- (i) 接触に対する反応(Thigmotaxis)。
- (ii) 化学物質に対する反応(走化性)。
- (iii) 温度に対する反応(Thermotaxis)。
- (iv) 光に対する反応(走光性)。
- (v) 電流に対する反応(ガルバノタキシス)。
- (vii) 重力に対する反応(地軸性)。
- ゾウリムシの生殖。
- (i)横方向の二体核分裂。
- (ii) 抱合:
- ゾウリムシの生殖における異常行動:
- (i) エンドミクシス。
- (ii) Autogamy:
- (iii) Hemixis:
- (iv) Cytogamy。
- Some Cytoplasmic Particles Reported in Paramecium Caudatum:
- (i) Kappa Particles.(カッパ粒子)。
- (ii) mμ、粒子。
- (iii) π粒子。
- (iv) ラムダ粒子。
生息地:ゾウリムシの細胞質微粒子報告
Habit, Paramecium Caudatumの生息地と培養。
Paramecium caudatum(Gr., 淡水の池、プール、溝、小川、湖、貯水池、川などに生息しています。 特に、腐敗物の多い淀んだ池や、有機物の注入口、下水などに多く生息している。 本種は自由生活生物であり、世界中に分布している。
ゾウリムシの培養。
ADVERTISEMENTS:
池から沈んだ雑草を取り、蒸留水の入った瓶に入れ、蓋をして腐らせておくと、数日後にゾウリムシの大群が現れます。 また、干し草を水で煮て、煎じ汁を捨て、数粒の小麦を加え、バクテリアで濁るまで放置する。
最初の瓶からゾウリムシをこの液体に移し、急速に増殖させる。 花粉の注入だけではシストを形成するゾウリムシが発生し、砂粒に似たシストが報告されているが、ゾウリムシがシストを形成することは確認されていないため、確証はない。
ゾウリムシの構造:
(i) ゾウリムシの大きさと形:
ミクロの生物で、肉眼では細長い体をしているのが見える。
体長は170〜290ミクロンで、薄い灰色か白色に見えますが、300〜350ミクロンになることもあります。 スリッパや靴の底のように見えるので、スリッパアニマルとも呼ばれる。 長さは幅の4倍で、やや円筒形で両端が明瞭に異なる。
前方の部分は細長く、先端は鈍いか丸みを帯びており、後方の部分はやや尖った円錐形をしています。 生物の最も幅の広い部分は、ちょうど真ん中より下にある。 体形は左右非対称で、口腔側と背中側が明瞭に区別される。
(ii)ペリクル。
体はゼラチンからなる薄く二重の弾力性のある固いペリクルに覆われています。 ペリクルは動物の形を保ちつつ、収縮できるような弾力性を持っている。 ペリクルは二重膜になっており、外膜は繊毛と、内膜はエクトプラズムと連続している。 高倍率の顕微鏡で見ると、ペリクルの表面には長方形や六角形の凹みがあることがわかる。

背面ではこの配列が残っているが腹面では前口径と後口径に向かって前後に隆起して収斂している。 六角形の窪みには中央に開口部があり、そこから1本の繊毛が出ている。 六角形のくぼみの前縁と後縁には、三葉虫の開口部がある。
EhretとPowersによるペリクル(図20.2)の電子顕微鏡的研究により、六角形のくぼみは規則的な一連の空洞である肺胞に相当することが明らかになった。 すべての肺胞は集合的に連続した肺胞層を形成し、この層は外肺胞膜と内肺胞膜で区切られている。
外膜は細胞外膜の下に密着している。 従って、ペリクルは外細胞膜、外肺胞膜、内肺胞膜を含む。
(iii) 繊毛。
体全体が繊毛と呼ばれる無数の小さな毛のような突起で覆われている。 繊毛は体全体に縦列しているが、このように体内の繊毛が均等な状態をホロトリキウスという。 繊毛は鞭毛と同じ構造をしており、外側の原形質鞘または形質膜の周辺に9本の二重縦線繊維が環状に存在している。 いくつかの繊毛では、9本の外側のフィブリルは対になっていない。
注意事項:
中央には2本の縦線があり、外側の縦線より細い。 各繊維は基底顆粒またはキネトゾームから生じている。 9対の周辺線維が融合してキネトゾームの壁を形成するため,キネトゾームは下端が開閉する管であり,ほとんどの繊毛虫では2本の中心線維はペリクルの高さで止まっている。
動原体から細い根粒が伸びているが、核とは結合していない。 多くのメタゾアにも繊毛があり、その構造は基底顆が異なり、細胞質内に伸びる細いフィラメントや根出繊維を持つ以外は同じである。 しかし、繊毛は一般に数が多く、大きさも短いという点で鞭毛と異なる。
繊毛は、体表にある体毛と、口の部分にある口腔繊毛とに分けられる。 体毛は等しいが、後端が長いので、caudatumと呼ばれる。 繊毛は運動や食物収集の器官であると同時に、感覚受容器として働き、外部環境の刺激を感知している。
繊毛の超構造。
繊毛と鞭毛は繊維状に構成されています。 繊毛は基部で約0.2ミクロンあるいは2000A0の直径を持ち、細胞表面から最大10ミクロンになることもある。 繊毛は厚さ90μmの単位膜に囲まれており、この膜は細胞膜に類似しており、連続したままである。 繊毛の束縛された空間には、マトリックスと呼ばれる水状の物質が含まれている。
マトリックスには、11本の縦方向のフィブリルまたは微小管が埋め込まれたままである。 11本のフィブリルのうち、2本は中央に位置し、残りの9本は中央のフィブリルの周囲に周辺的に配置されたままである。 9本の外側のフィブリルはそれぞれ直径360Aで、直径180〜250Aの2本のサブフィブリルから構成されている。
これらのサブフィブリルをサブフィブリルAおよびサブフィブリルBとする。
注意:
A線維はB線維より少し大きく、片側から2本の太い突起(腕)が出ています。 すべてのアウターフィブリルのサブフィブリルAの腕は時計回り方向を保っている。 さらに、サブフィブリルAはサブフィブリルBよりも繊毛の中心により近い位置にある。
中央の2本のフィブリルには、周辺の9本のフィブリルのように対になったサブフィブリルはなく、それぞれ1本の細管のみが存在する。 各中央フィブリルの直径は約250A0であり、厚さ60A0の壁で構成されている。
両中心線維は350A0の空間を隔てて、共通の鞘に包まれたままである。 Gibbnos (1967)は中心線維の鞘が各サブフィブリルAに放射状に9本のリンクまたはスポークを出すと報告している。

高分解能電子顕微鏡により、繊毛と鞭毛の周辺線維と中心線維はそれぞれ40Aの太さで10〜12本の繊維からなることが明らかにされた。 各フィラメントはビーズ状である。 各ビーズは、管壁の平面上に40×50A0の格子状に配列したままである。 このビーズが細管構造の基本的なサブユニットと考えられている。
(iv)毛細血管内系。
毛細管内系は、小胞状肺胞のすぐ下に位置しています。 キネトゾームまたは基底体とキネトデスマからなる。 繊毛はキネトソームから発生し、それぞれのキネトソームからキネトデスマと呼ばれる繊細な細胞質繊維が発生する(Fig.20.2)。
ペリクルの下、やや右側に横たわっているが、1つの縦列のすべてのキネトゾームに結合しているのは、いくつかのキネトデズマの縦束で、各キネトゾームのキネトデズマは、前方にそれ自身のキネトデズマ束に距離をおいて伸びている。
キネトソームの縦列とそれらのキネトデスマータは、キネティと呼ばれる縦列単位を形成する。 すべてのキネティまたはキネティアは繊毛虫の繊毛内系を構成する。 キネティはペリクルの下の皮質にあり、その数は各繊毛虫でほぼ一定である。
繊毛内膜系は繊毛の動きを制御・調整し、細胞分裂の際に小器官の形成をもたらすが、例えば、いくつかの繊毛は口を形成する。 繊毛虫の2回分裂では、繊毛が横方向に2つに切断され、それぞれが1つの娘細胞に行くが、これはペリキネタルフィッションと呼ばれる。
(v)口腔溝とサイトピージ。
腹側には、口腔溝または口輪と呼ばれる大きな斜めの浅い窪みがあり、これが動物を非対称に見せている。 片側から斜め後方(通常は左から右だが、右から左の場合もある)に走り、中胸の少し後ろで終わっている。 口腔溝は前庭と呼ばれる短い円錐形の漏斗状の窪みに通じている。
前庭は直接、サイトストーム(口)と呼ばれる楕円形の固定された開口部につながっています。 細胞腔から体の中心に向かって直接伸びているのが、広い細胞咽頭です。 細胞咽頭はその後、後方に向かって急に曲がり、細長い食道となる。
このように食道は、後端を除いてゾウリムシの体表とほぼ平行になっています。 ここで食道は再び動物の中心に向かって曲がり、形成中の食胞につながる。
細胞肛門(cell anusまたはanal spotまたはcytoproctとも呼ばれる)は、細胞胞または口の後ろのほぼ垂直に体の腹面にある。 未消化の食物は、細胞吻から排出される。 細胞咽頭の繊毛は非常に複雑である。
Gelei(1934)は4列あると報告し,Lund(1941)は少なくとも4列を観察している。 細胞咽頭の左壁にはpenniculusと呼ばれる構造があり,約90度螺旋を描いて後端が食道の口腔(腹側)面に位置する。
Lundによると、小胞体は8列の繊毛がそれぞれ2つのブロックに密接して配列されている。 また、penniculusよりもコンパクトな4列の長い繊毛からなる同様の帯は、quadrulusと呼ばれる。 この帯は頬腔背側壁を螺旋状に下り、半月板近くで終わる。
半月板と四角板を誤って “undulating membrane “と呼ぶ研究者がいる。 この膜は食物の通過を制御している。 繊毛の働きは不明であるが、おそらく繊毛の繊維がリズミカルに収縮して曲がるのであろう。 Gelei (1925)は、penniculusの機能が食物成分の体内への押し出しであることを指摘している。

(vi) 細胞質:
細胞質は、外部または皮質の狭いゾーン(エクトプラズム)と内部または髄質の大きな領域(エンドプラズム)に区別されます。
(vii) Ectoplasm:
Ectoplasm (ectosarc or cortex) は体の永久部分で、内部質から顕著に区切られます。 エクトプラズムは堅固で透明な、薄くて密な外層を形成している。 三葉虫、繊毛、フィブリル構造を含み、ペリクルと呼ばれる被覆で外部に境界を接している。
(viii) トリコシスト(Trichocysts)。
エクトプラズムの中に、表面と直角に、トリコシストと呼ばれる小さな紡錘形の袋が埋め込まれています。 六角形の前縁と後縁にそれぞれ小さな点があり、これがトリコシストの位置を示している。 これらは膨潤物質を持つ屈折した濃厚な液体で満たされており、外側の端には円錐形の頭部またはトゲがある。
三胞子はエクトプラズムの中で垂直に並び、ペリクルの六角形の部分の稜線にある小さな孔で開きます。
繊毛のキネトゾームから発生し、移動して小胞体内で等間隔に位置する。 動物が刺激を受けると、三毛子嚢は長い粘着性の糸となって排出される。 排出されたトリコシストには、逆さ爪のような不透明なトゲと、長い筋状の軸があるが、軸は未排出の状態では見られず、排出時に形成されると思われる。
トリコシストの機能は不明であるが、局所的な接触や傷害に対する反応として排出されることから、防御器官として機能している可能性がある。
しかし、ゾウリムシの主な捕食者であるDidiniumには効果がないので、これは不確かで、摂食時に動物をある場所に固定するためのものかもしれない。 また、繊毛虫のなかには、三叉球が攻撃器官として働いているものもある。 また、キネトゾームから再生されたトリコシストが排出される。


(ix) 神経原線維と関連原線維(Paramecium and Associated Fibrils)
Lund(1933)によると、細胞咽頭の左背部壁で細胞突起の後縁の高さに、非常に小さな二枚貝の塊、神経運動器がある。 この神経原器から、線維が放射状に伸びてエンドプラズムに入る。
これらのうち、4本以上は通常ほぼ背側の体壁まで達していますが、残りは短く、位置も定かではありません。 これらはすべて小胞体線維と呼ばれる。 その機能は不明であるが、口腔繊毛の摂食運動を調整しているのかもしれない。 また、フィブリルは、機械的支持、弾性、収縮性、伝導性、代謝の影響を与えることがあります。

(x) 内膜:
内膜または髄質は、細胞質のより流動的で多量の部分で、多くの細胞質顆粒と他の包有物と特殊な性質の構造物が含まれています。 細胞質内の包有物はミトコンドリア、ゴルジ装置、液胞、結晶、顆粒、染色体などである。 その他の構造、すなわち核、収縮液胞、食液胞も小胞体内に存在する。
核。
細胞質近くの小胞体には2つの核、すなわちゾウリムシは異核性で、大きな楕円形と粒状の大核と、小さなコンパクトな小核がある。
巨核は楕円形か腎臓形の顕著な体型である。 大きさの異なる細糸状の不連続なクロマチン顆粒を含み、無色のマトリックスに埋め込まれたコンパクトなタイプである。 核小体も多く、クロマチン物質(DNA)も非常に多い。
体細胞核または植物核である。 単調に分裂し、動物の植物的な機能(代謝活動)を司る。 有糸分裂を行う。
小核は小さく、コンパクトで球形をしています。 一般に大核の近くにあり、凹んでいることが多い。 微細なクロマチン顆粒と糸が構造全体に一様に分布している。 この微小核は分裂を行い、生殖を司る。
微小核の数は種によって異なり,P. caudatumでは1個,P. aureliaでは2個,P. multimicronucleatumでは多数である。 微小核はP. aureliaでは明瞭な核小体を含むが,P. caudatumでは見られない。 Moses (1949; 1950) は,大核と小核の化学組成が同一であることを報告している。
(xi) 収縮性液胞:
体の片端から背面近くに2つの大きな液胞があり,それぞれ液体で満たされる。 その位置は(アメーバと違って)一定で、外形質と内形質の間に位置するが、周期的に消失する一時的な小器官である。 種によっては裏打ち膜があるようで、その場合は収縮期に完全に消滅することはない。
各収縮液胞には5〜12本の管状の放射管があり、それぞれ末端部、空になるとつぶれる長いアンプル、液胞に開く短い注入管からなる。

管は体の大部分と連絡しており、そこから液体を取り込んで液胞に注ぎ、このように再構成されて大きくなり、収縮液胞がその最大サイズに達すると、突然収縮して(収縮期)その内容を小胞の永久孔を通して排出し、その後管はまた収縮液胞を形成するが、管は永久構造であるので完全に消滅することはない。
2つの収縮胞は不規則に排出されるが、後方の胞は細胞咽頭に近く、より多くの水が入ってくるので、より急速に収縮する。 水路と収縮液胞の主な機能は静水性で,原形質から過剰な水を除去し,水は摂食中に一部が吸収され,一部が取り込まれる。
窒素系排泄物にはアンモニア化合物といくつかの尿酸塩があり、これらはCO2と一緒に収縮液胞から排出されるが、原形質から運河や収縮液胞に排泄物質が分泌された形跡はない。
細胞咽頭の片側にある口蓋垂の下には、固定した永久細胞肛門があり、そこから未消化の食物の残骸や尿酸塩が排出される。
収縮胞の電子顕微鏡的研究により、各収縮装置は小胞体の管、腎臓の管、送り出し管、付属胞(放射状管)および主収縮胞の一部からなることが判明した。 付属液胞はフィーダー管の管腔であると推測される(図20.11)。
(xii)食液胞。
これらはほぼ球形の非収縮性体で、大きさも数もさまざまで、小胞体内に横たわっています。 この中には、摂取された食物粒子、主にバクテリアと、薄い膜に囲まれた少量の液体が含まれている。 Volkonsky (1934)はこれらの液胞をガストリオールと呼ぶことを提案した。 食物の液胞と関連しているのは消化顆粒である。
ゾウリムシの運動:
ゾウリムシは代謝や体の歪みと繊毛による2つの方法で運動する。
(i)メタボリックまたは体の歪み。
ゾウリムシの体は弾力性があり、体より狭い通路を通ることができ、その後、体は元の形に戻ります。 この一時的な体の形の変化は代謝であり、ゾウリムシでは原形質によってもたらされます。
(ii) Ciliary Locomotion(繊毛運動)。
繊毛によってもたらされるlocomotionが主な方法である。 繊毛は前方または後方に拍動することができ、動物は前方または後方に泳ぐことができる。
通常、動物は前方に泳ぎ、繊毛は後方に鼓動するが、斜めに、繊毛は硬くなり、体表面にほぼ触れるまで急速に後方に曲がる、これは有効ストロークと呼ばれる; 次に繊毛はぐったりして元の垂直位置にゆっくりと戻り、これは回復ストロークと呼ばれる。
同じ横列の繊毛は一緒に拍動し、同じ縦列の繊毛は前端から後端に向かって次々に拍動する。

この繊毛の協調運動はメタクロナルリズムと呼ばれ、毛内系に起因し、これにより動物は前方に泳ぐ。 しかし、体繊毛が斜め後方に拍動しているとき、同時に口腔溝の長い繊毛がより激しく拍動し、これにより前端が左に振れるのである。
体腔溝と口腔溝の繊毛の働きによって、動物は長軸方向に回転することになる。 この回転は常に左回りである(ただし、P. calkinsiは右回りの螺旋状に回転する)。
この前進、旋回、回転の組み合わせにより、反時計回りの螺旋軌道を描いて前進する。 この経路は軸が直線であり、動物の同じ体表は螺旋経路の軸の方に残る。 しかし、後方へ泳ぐ際には、すべての種が右回りに回転する。
繊毛の拍動を逆にして、繊毛を斜め前方に移動させ、それによって動物は後方に泳ぐことができる。 毛細管現象により、ゾウリムシは1秒間に1500ミクロン、あるいはそれ以上の速度で移動する。
ジェニングスは、ゾウリムシが螺旋状に動くのは、繊毛が主に後方に打ちながら、右斜めに打ち、それによって動物を左側に横転させるからであると主張した。
また、このように体が口腔面に向かって旋回するのは、より直接的に後方に衝突する口腔繊毛の有効ストロークの力が大きいことが大きな原因である。 その結果、ゾウリムシはその長軸を中心に回転し、大きな渦巻きを形成する際に、多かれ少なかれ直線的な経路をたどることができるようになりました。
Paramecium Caudatumの栄養:
Paramecium Caudatumでは、栄養は全生代的である。 餌は主にバクテリアと微小な原生動物です。 餌を待つのではなく、積極的に狩りをする。
餌の選択に選択の余地があると主張されるが、特定の種類の細菌しか食べず、大腸菌を24時間に1匹のゾウリムシが200万~500万個食べるというデータがあることから、根拠はないようである。 また、藻類や珪藻類などの単細胞植物や、動物や野菜の小片も食べる。
摂食のしくみ。
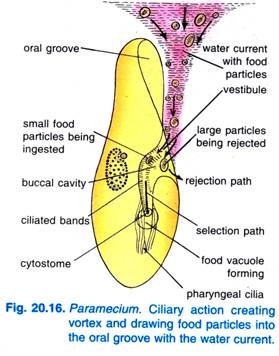
Paramecium Caudatumは、餌が豊富な地域に入ると、休息に入ります。 餌は静止しているときか、非常にゆっくり泳いでいるときだけで、速く泳いでいるときには決して食べません。 口腔溝の繊毛の拍動によって、餌を含んだ円錐状の水の渦が前端より前方から口腔溝内に流れ込む(図20.16)。
その後、食物の粒子は前庭に向かい、そこから一部の食物の粒子は拒絶されて投げ出されるが、他のものは細胞質へと通過していく。
細胞咽頭の末端には食物胞が形成され、食物の粒子で満たされる。 四徴と小腸は側方に形成された食胞への食物の通過を制御している。 食胞がある大きさになると、後頬骨繊維が食胞をはさみ、食胞はこれによってつまみ出され、その軌道を進み始める。
食胞の中には食物のほかにいくらか水分が含まれている。 サイクロシスと呼ばれるエンドプラズムの回転運動が食胞を一定のコースで運ぶが、これは機能的には消化管に相当する。
管は細胞咽頭の端から始まり、次に後方へ、そして前方でエンドプラズムと循環し、次に背面へ、そして前端へ、そして下方の細胞辺縁へ向かう。 食胞は移動の初期にその大きさが減少し、その後再び増大する。

Paramecium Caudatumの消化と排泄:
サイクロシス中に、原形質から小胞体に分泌される酵素によって消化が行われる。 消化の過程で、タンパク質はアミノ酸に、炭水化物は可溶性糖とグリコーゲンに、脂肪もおそらく消化される。
食物の液胞の内容物は最初酸性(pH約4)で、その後アルカリ性になり、主要な消化はアルカリ性の段階で行われる。 未消化物はある程度の力をもって細胞胞から排出される。
サイクロシスは実験的に示すことができ、コンゴレッドで染色した牛乳をゾウリムシに与えると、食物胞内の牛乳の脂肪球は酵素の酸性反応によりまず赤くなり、次にアルカリ反応により紫から青へと変わり、胞巣はサイクロシスの経過を示すようになります。
ゾウリムシの呼吸と排泄:
ガス(酸素と二酸化炭素)の交換は、他の淡水原生動物と同様に半透膜を通して、拡散の過程で行われます。 ゾウリムシは周囲の水から酸素を得る。 代謝によって生じた二酸化炭素やアンモニアなどの有機性廃棄物は、逆方向の水中へ拡散して排泄されるのであろう。

6. ゾウリムシの体液調節:
ゾウリムシには前部と後部の2つの収縮液胞がある。 収縮液胞の機能は体内の水分調節であり,尿素やアンモニアなどの窒素系老廃物の排泄も担っている可能性がある。
細胞質内の過剰な水(連続的なエンドースモシスによる)は小胞体の管に分泌され、腎臓の管→送り出し管→へ進み、6〜11本の放射状の管が各液胞に向かって収束し、その中に排出される一連の管に集められる。 この管は液胞が形成されるときに最も顕著に現れる。
それぞれの液胞がある大きさに膨らむと(拡張期)、収縮して(収縮期)、おそらく気孔を通して外部に排出される。 収縮液胞は10〜20秒の間隔で、交互に収縮する。
後方の収縮液胞は、細胞咽頭から大量の水を後方に取り込むため、前方の液胞より速く働く。 収縮液胞は余分なものを処理することで体細胞質内の水分濃度を最適な状態に保つ。

Paramecium Caudatumの行動:
様々な種類の刺激に対するゾウリムシの反応は、その反応や培養中の個体の集団化や散乱を調べることによって知ることができる。 反応は刺激に向かうと正になり、遠ざかると負になる。 逆刺激に対しては、動物は逃げるまで回避反応をし続ける。
回避反応では毛様体拍動が反転し、少し後方に移動した後、後端を軸に前端を外側に振って円錐軌道を描くように回転する。 すべての調整は試行錯誤で行われる。 実験によると、前端は他の部分より敏感であることが分かっている。
異なる刺激に対するゾウリムシの反応は、次のようにグループ分けできる。
(i) 接触に対する反応(Thigmotaxis)。
ゾウリムシでは接触に対する反応はさまざまです。 前端を細いもので軽く触ると、強い回避反応が起こる。 泳いでいるゾウリムシが水中で何かの物体に衝突しても、他の場所を触ると無反応になることもあります。 動きの遅い個体は、物体に接触すると積極的に反応して、その上に静止することが多い。
(ii) 化学物質に対する反応(走化性)。
一般にゾウリムシは化学的刺激に対して、反応を避けるという方法で反応します。 マイクロスライド上のゾウリムシの集団に0.5パーセントの弱い塩溶液を一滴たらすと、回避反応によって反応し、一滴も中に入らなくなる。 しかし、酸に対しては、死滅させるのに十分な強さの濃度であっても、反応は陽性である。
(iii) 温度に対する反応(Thermotaxis)。
ゾウリムシは24~28℃を至適温度とする。 最適温度の上下で著しく温度が変化すると、ゾウリムシは回避反応を示す。 熱量が大きくなると、動物が逃げるか殺されるまで、急速な動きと回避反応が促される。
(iv) 光に対する反応(走光性)。
積極的な光走性を持つ緑色ゾウリムシを除いて、他の種は普通の光に無関心である。 しかし、光強度を突然急激に増加させると、一般に負の反応が続く。 紫外線に対しては、ゾウリムシは直ちに負の反応を示す。
(v) 電流に対する反応(ガルバノタキシス)。
ゾウリムシは電気刺激に反応します。 ゾウリムシの入った浅い皿に2つの電極を対向させて一定の電流を流すと、すべての生物は陰極またはマイナスの電極に向かって同じ方向に泳ぎ、そこに大量に集中するようになります。
ゾウリムシが陰極に向かって泳いでいるときに電流の方向を逆にすると、生物は方向を変えて新しい陰極に向かって泳ぎます。

⑥水流に対する反応(Rheotaxis)。
ゾウリムシは正のレオタクシスを示す。 緩やかな水流では、ゾウリムシはほとんど流れに乗り、前端を上流に移動する。
(vii) 重力に対する反応(地軸性)。
パラミシアは一般に重力に対して負の反応を示し、多くの個体が前端を上にして表面膜の下に集まっているのが観察されます。 また、水を張った逆さU字管にゾウリムシを入れると、すぐに水平部分に移動してしまう。
ゾウリムシの生殖。
Paramecium Caudatumは横二核分裂によって無性生殖を行い、またコンジュゲーション、エンドミキシス、オートガミー、サイトガミー、ヘミキシスなどいくつかのタイプの核再形成を行う。
(i)横方向の二体核分裂。
横型二体核分裂はゾウリムシの無性生殖の中で最も一般的なタイプである。 これは、1つの完全に成長した標本が、親の死体を残さずに2つの娘個体に分裂するという、明らかにユニークな無性生殖過程である。
分裂の面は細胞の中心を通り、体の長軸に直角な面である。 細胞体全体としての分裂は常に核の分裂に先行し、実際、生殖は核の活動と分裂によって開始されるようである。

Paramecium Caudatumは好ましい条件下では横方向の二回核分裂によって生殖する。 二元分裂では、微小核が有糸分裂によって2つの娘微小核に分裂し、細胞の反対側の端に移動する。 大核は伸長し、アミトーシスによって横方向に分裂する。
別の細胞鞘が出芽し、前端付近と後端付近の2つの新しい収縮液胞が出現する。 一方、中央付近には狭窄溝が現れ、細胞質が完全に分裂するまで深くなっていく。
こうしてできた2つの「娘」ゾウリムシは同じ大きさで、それぞれが細胞小器官を一式持っています。 生成された2つの娘ゾウリムシのうち、前方のものはプロター、後方のものはオピステと呼ばれる。 この2つの娘ゾウリムシは、次の分裂が起こる前に完全に成長する。
二元分裂の過程は約2時間を要し、1日に1~4回起こり、2~16個の個体ができる。 1年間に約600世代が生産される。
増殖の速度は、食物、温度、培養の年齢、人口密度などの外的条件と、遺伝と生理の内的要因に左右される。 当然ながら、1個体の子孫がすべて生存して繁殖すれば、生産されるゾウリムシの数はすぐに地球の体積に匹敵することになる。
1つの個体から核分裂によって生じたすべての個体を指してクローンという言葉が使われる。 クローンの構成員はすべて同じ遺伝子を持つ。
(ii) 抱合:
通常、ゾウリムシは二元分裂で長期間増殖するが、ある時期に2匹が口腔面に沿って結合し、抱合という性行為で中断されることがある。
コンジュゲーションとは、微小核物質を交換し合う2つの個体の一時的な結合と定義される。 核物質を交換した後、すぐに分離する特殊な性行為である。
Sonneborn (1947) は、ゾウリムシの交尾行動に基づいて、ゾウリムシの各種は多数の品種または合性で存在すると報告しています。 さらに、それぞれのsyngenの中には、通常2つの交尾型が存在する。
これらの交配型は形態的には同じであるが、生理的な違いが見られる。 P. aureliaでは14個のsyngenと28個のmating typeがあり、P. caudatumでは16個のsyngenと32個のmating typeがある。 また、通常ゾウリムシは自分の交尾型とも他の交尾型とも交尾せず、自分の交尾型の第2交尾型としか交尾しないことが観察されている。

共役を誘発する因子。
共役を誘発する因子は種によって異なるが、そのいくつかを以下に挙げる。
1. 飢餓や餌の不足、特定の細菌の餌や化学物質が、ある種のゾウリムシの抱合プロセスを誘発すると言われている。
2.抱合は約300回の無性核分裂を経て、あるいは長い間隔で無性核分裂と交代して瀕死のクローンを若返らせる、つまり未熟期といわれる望ましい数の無性世代を経て、性成熟して抱合する個体に発生する。
3.抱合はゾウリムシの生理状態に変化が生じたとき、体長が210ミクロンとやや小さく、不健康な老齢期にある個体間で起こり、抱合させなければ死んでしまう。
4.突然の暗黒や低温が抱卵を誘発する種もあるといわれる。
5. 夜間や暗闇の中では抱合は起こらず、早朝から午後まで抱合は続く。
6.交配型個体の繊毛に含まれるタンパク質性物質が抱合を誘発すると言われている。
抱合過程。
抱合過程はゾウリムシの種類によって異なるが、ここではP. caudatumの抱合過程について記す(図20.21)。
抱合では、同じ品種の反対の交尾型の2つのゾウリムシ(前抱合体という)が腹面を合わせて、口腔溝で結合し、その繊毛から体表に物質を出して、2つの抱合ゾウリムシを付着させるのです。
摂食を停止し、口腔溝器は消失する。 両者の接触点であるペリクルとエクトプラズムが破壊され、2匹の間に原形質の橋が形成される。 さて、これらの個体はコンジュガントと呼ばれる。
この状態で、抱卵対は活発に泳ぎ、同時に各抱卵体には以下に述べるような一連の核変化が起こります。
大核は崩壊を始め、緩い質感と複雑な撚り糸を形成し、抱合期後半には細胞質内に吸収されて消滅する。 各結合体の小核は2回分裂し、そのうちの1回は還元分裂をする。
このようにして、各結合体には4個のハプロイドの娘微核が生じる。 この4個の微小核のうち3個はそれぞれ退化し、1個だけが残る。
各結合体の残りの微小核は、分裂して2つの不等な前核または配偶子核になり、より大きな静止雌前核とより小さく活発な移動性雄前核を形成します。
一方の前核は原形質橋を渡り、他方の前核と融合し、染色体の2倍体が復元され、遺伝物質の交換が行われた合核を形成します。
この過程は高等動物の受精と比較されるが、配偶子が関与しないので受精とはいえない。 このとき(約12~48時間後)、結合体は分離し、元結合体と呼ばれる。 各前成体のシンカリオンは3回分裂して、各前成体に8個の微小核を形成する。
8個の微小核のうち4個は大きくなって巨核となり、残りの4個の微小核のうち3個は消滅する。
残りの微小核が分裂すると同時に、元結合体は二核分裂して、それぞれが2つの巨核と1つの微小核を持つ2つの細胞になる。 この細胞および微小核は2回目の分裂を行い、それぞれの元結合体から4匹のゾウリムシを形成し、それぞれが1個のマクロ核と1個の微小核を持つようになる。
新しい大核は、小核と同様に新しい材料でできています。 これらの新しい核は、おそらく新しい別の潜在能力を含んでおり、それが健康な個体に反映されているのであろう。
結合の意義。
核の再編成が行われないとクローンは死滅するが、核の再編成によってクローンは以前の活力を取り戻すことができる。この核の再編成は共役によってもたらされ、したがって共役は二元分裂を続けるために不可欠である。
共役の意義については、以下にまとめた。
1. 結合は若返りと再組織のプロセスとして機能し、それによって種族の活力が回復される。 長期間抱合しない場合、ゾウリムシは弱り、死んでしまう。 (ウッドラフは2万2千世代にわたってゾウリムシを健全な状態に保ったと主張しているが、ゾンネボーンはウッドラフのゾウリムシがすべて同じ交配型に属していることを示し、これを否定している)。
2.同じ品種の異なる2つの交配型のゾウリムシだけが交配するが、交配体には性別の区別がない。
3.雌雄の区別はないが、活発に移動する前核を雄、静止している前核を雌と見なした。
4.抱合は一時的な結合に過ぎず、細胞質の融合はなく、接合子は生じないが、各前抱合子の核は2つの抱合個体の遺伝物質を含んでいる。
5. 共役は大核を共核からの物質で置き換えるが、これは基本的に重要な出来事である。 二核分裂では大核の染色体はランダムに娘細胞に分配され、二核分裂を続けることでクローンは弱くなり、いくつかの構造的異常が生じた。
共役により、大核に正しい数の染色体が形成され、種族が新しく元気に生まれ変わる。 小核の役割は、バランスのとれた染色体と遺伝子の複合体を回復させることである。
ゾウリムシの生殖における異常行動:
ゾウリムシは分裂と抱合時の核の挙動に一定の変化を示し、これらの逸脱はエンドミクシス、オートガミー、サイトガミーおよびヘミクシスである。 最初の3つの過程では、遺伝子組み換えが起こり、微小核から新しい大核が形成される。
(i) エンドミクシス。
Woodruff and Erdmann (1914)はまず、2核生物であるParamecium aureliaで新しい核再組織プロセス、エンドミクシス(Gr., endon = within; mixis = mingling)を報告している(図20.22)。 この過程は、周期的に起こり、シンカリオン形成なしに新しい大核装置が作られると説明された。 エンドミクシス(endomixis)は1つの個体内で起こる。
WoodruffとErdmannによると、大核は退化し、小核は2回分裂して8個の小核を形成する。 6個の微小核が退化し、2個が残る。 2個の微小核が残った状態で、核分裂により2つの細胞に分裂し、それぞれが1個の微小核を持つようになる。
各娘細胞の微小核は2回分裂して4個の微小核を形成します。 2つの微小核が大きくなって巨核になります。 動物とその微小核が分裂して、それぞれ1つの巨核と2つの微小核を持つ2つの娘個体が形成されます。
エンドミキシングは抱合しないP. aureliaの品種でも起こる。したがって、エンドミキシングの効果は抱合の効果と同じかもしれない。どちらのプロセスもマクロ核をミクロ核の物質で置き換え、レースの生命力を若返らせるからだ。
しかし、内殖では前核の融合がないため、この2つの過程は異なっており、内殖は単為生殖と比較することができる。
しかし、一部の研究者は、内生殖は有効な過程ではなく、誤った観察により記述されたものであると、正当な理由を持って主張している。 しかし、エンドミキシングは有効なプロセスではなく、誤った観察により記述されたものであると主張する研究者もいる。

その後、Erdmamm and Woodruff (1916) はParamecium caudatumでendomixisを報告している。 しかし,Dillerはこの過程の妥当性を信じておらず,ErdmannとWoodruffはhemixisとautogamyの段階をendomixisという一つの方式に統合しただけだと考えている。
(ii) Autogamy:
Diller (1934, 1936)とSonneborn (1950)は、ゾウリムシの単一個体で起こる自己受精、すなわちオートガミーについて述べた(Fig. 20.23)。 彼は、自家受精では、成熟を伴う3回の微小核(前核)分裂によって配偶子核(前核)が生成されると報告している。
オーレリアの自殖では、2つの微小核が2回(減数分裂1回)分裂して8つの微小核を形成し、そのうち6つは退化する。 一方、大核は綛状に成長し、後に分裂して細胞質内に吸収される。 8個の微小核のうち2個は前核として、細胞口付近で膨らんだ原形質コーンに入る。
2つの前核は融合して合核を形成します。 シンカリオンは2回分裂して4個の微小核を形成する。 2個の微小核は巨核となる。 ゾウリムシとその微小核が分裂して、1つの巨核と2つの微小核を持つ2つの娘個体ができる。 この過程は2日程度で完了する。
Autogamyは種族の若返りをもたらすのです。
微小核の材料で新しい大核が形成され、新しい大核では染色体の数が正しく回復する点、また2つの前核の融合が起こる点で、受精と類似しています。 しかし、自家受精は1つの個体だけが前核を提供し、一種の自己受精であるため、結合と異なる。
(iii) Hemixis:
Diller (1936)はParamecium aureliaで半数分裂を報告した(Fig. 20.24)。 半数分裂は主として大核の断片化と分裂の過程であり、微小核の異常な活動はない。 Dillerは、P. aureliaで図20.24に示したように、hemixisをA、B、C、Dの4つのタイプに分類したが、P. caudatumやP. multimicronucleatumの大量培養でもすべてのタイプに遭遇している。
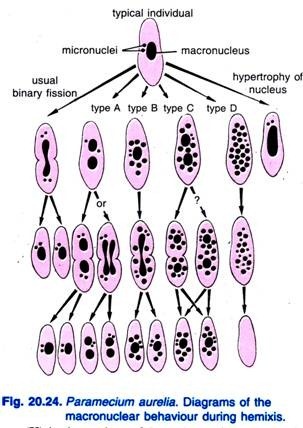
A型は大核が二つ以上に分裂する最も単純な半数分裂の形態であり、その特徴は大核が二つ以上に分裂していることである。 この分裂は微小核の分裂と同期していない。
B型は、1個から20個以上のクロマチンボールが巨核から細胞質へ押し出されることが特徴である。
C型は、大核が2つ以上の主要な部分に同時に分裂し、大核球が細胞質へ押し出されることが特徴である。
D型は、大核がクロマチン球に完全に分裂し、最終的に細胞から消失する病的状態を表すと考えられている。 小核は一般に大核が溶解する前に消失する。
(iv) Cytogamy。
Wichterman (1939)はParamecium caudatumの別の性行為を報告し、彼はこれをcytogamyと名づけた。 細胞婚では核の交換はない。 この場合,2つの個体は腹面によって結合するが,2つの個体のペリクルは破壊されない。
各個体の小核は3回分裂して8個の小核を形成し、そのうち6個は各個体で分解する。 残りの2個の微小核は融合して、各細胞の中でシンカリオンを形成する。 これで動物が分離する。
細胞結合は、2つの動物が接触している点では自家発電と異なるが、2つの前核が融合する点では自家発電や抱合に類似している。 Cytogamyは,接触する2つの動物の間で核の交換がない点でconjugationと異なる。
Some Cytoplasmic Particles Reported in Paramecium Caudatum:
(i) Kappa Particles.(カッパ粒子)。
1938年、T.M. Sonnebornはゾウリムシのある種(キラー株と呼ばれる)がパラメシンと呼ばれる毒物を作り、これが感受性のある他の個体に致死的であると報告しました。 このパラメシンは水溶性で拡散性があり、ゾウリムシ(キラー株)の細胞質内にあるいくつかの粒子に依存して生成される。
この粒子はカッパ粒子と呼ばれています。 カッパ粒子はDNAとRNAを持っている。 キラーゾウリムシは何百ものカッパ粒子を含んでいることがある。 この粒子の詳細な研究により、カッパ粒子が存在し、増殖し、パラメシンを生産するためには、ゾウリムシの核にある優性遺伝子(K)が必要であることが判明した。
(ii) mμ、粒子。
R.W.Siegel (1952)はいくつかのゾウリムシの細胞質中にもう一つのタイプのキラー粒子を報告した。 mp粒子を持つゾウリムシは、mate sensitiveと呼ばれるmμ粒子を持たないゾウリムシと結合すると、後者を殺すので、mate killerと呼ばれる。 mp粒子は、DNAやRNAなどからも構成されている。
これらの粒子は、連鎖していない2対の染色体遺伝子(M1およびM2)のうち、少なくともどちらかの優勢遺伝子を小核に含むゾウリムシにのみ存在する。
(iii) π粒子。
この粒子はカッパ粒子の変異型と思われるが、いかなる種類の毒物も生成しない。
(iv) ラムダ粒子。
この粒子は殺人ゾウリムシの中に報告され、これを持たない敏感なゾウリムシを溶解または崩壊させる何らかの物質を生産すると言われています。