ボルボックスの事例(分類、特徴、顕微鏡下のボルボックス、繁殖(無性生殖と有性生殖)
ボルボックスは淡水の浮遊性藻類です。 この属には20種ほどが属しています。 植物界で最も美しく、魅力的な存在として知られています。 淡水域ではピンの頭ほどの大きさの緑色の球体で見られる。
Detailed Guide With All Aspects
藻は、特定の泳ぐパターンがあるため、しばしば、ローリング藻と呼ばれています。 その成長は、一時的または恒久的な淡水の池、プール、溝、また湖で頻繁に観察されます。 水温と有機物が十分な量ある場合に多く繁殖する。 池の水が緑色になる(ウォーターブルーム)ほど、頻繁に、そして大量に増殖する。 春と雨の季節は、通常、植物が活発に成長する時期である。 不順な季節(夏)になると藻は消え、接合体の形で不順な季節を過ごす。 Volvoxは光合成によって彼ら自身の食糧を作る。
Volvox分類
- Class: 緑藻類
- オーダー。 Volvocales
- Sub-order: Chlamydomonadineae
- Family: スフェレラ科
- Genus: Volvox
Volvoxの最も一般的な種はV, globator, V, aureus; V. prolificus, V. rouseletti, V. merelliである.
ボルボックスの特徴
植物体(配偶体)
ボルボックスは共生の緑藻類で、{(コロニー-植物体は細胞の数が一定でない例:ボルボックス・オーレウス)(共生-植物体は細胞の数が一定、例:…ボルボックス・オーレウス)。 Pandorina morumaの場合、細胞数は4、8、16、32。 Eudorina unicocca、細胞数16、32または64)}。 運動する形態の中で、Volvoxのcoenobiumは最も大きく、高度に分化し、よく進化した藻類である。 各コエノビウムは楕円体または中空の球体で、繊細な粘液層が正確に記されている。
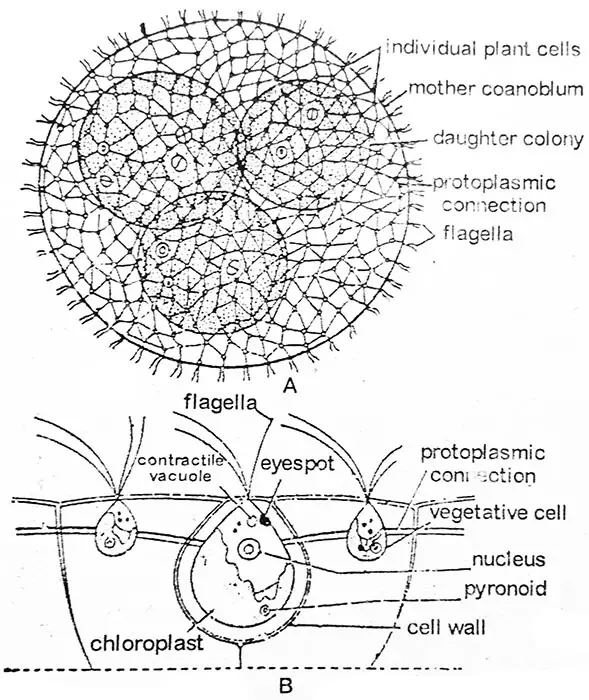
コロニーの運動は、個々の細胞の鞭の共同作用でもたらされる。 それぞれのコエノビウムには明確な前端と後端がある。 コエノビウムは極性を示し,ゆっくりと移動・回転し,その運動の過程で前端と後端の細胞間の顕著な協力関係を示している。 Volvoxは個体ではなく、多数の類似した細胞の連合体であり、それぞれが独立した個体のように機能し、独自の栄養、呼吸、排泄を行い、これらの機能において細胞間の協力は見られない。
細胞の構造
研究によると、個々の細胞はそれぞれ個性を持っている。 それはそれ自身の大きなゼラチン状の、鞘に囲まれており、隣接する細胞の鞘と相反する場合もあれば、互いに異なる場合もある。 その場合は、互いに圧縮されて角ばった形になり、外形は通常、六角形である。 図.2.22. このように、かなりの広さのゼラチン質が、一つの細胞を他の細胞から分離するのに役立っている。 多くの種では、各細胞は、細胞分裂やコロニーの形成の過程で形成された一連の原形質または細胞質ストランド(形質膜、図2.22)によって、隣接する細胞と連結されている。 原形質線は,V. aureusでは細く繊細で,V. globatorでは丸く,V. mononaeではくさび形であったり,V. tertiusのように存在しないこともある
Volvoxの個々の細胞の輪郭はChlamydomopasに似ている. 各細胞の前方には長さの等しい一対の鞭毛が挿入されている。 鞭毛は2本ともむち打ちタイプである。 鞭毛はコエノビウムの表面から周囲の水中に突出する。 鞭毛の基部付近には2個以上の収縮性液胞が存在する。 プロトプラストは種によって異なる形をしている。 V. aureusのプロトプラストは丸みを帯びたクラミドモナス型であるが,V. globatorのプロトプラストは星型で,拡散した葉緑体と散在した収縮液胞をもつ. この細胞は無数のピレノイドを形成して大きくなり、生殖細胞として機能する。 無性細胞である場合と有性細胞である場合がある。 同じコロニーに無性細胞と有性細胞の両方が存在することもある。 前部では、コロニーの細胞は植物的で比較的小さいままである。 前方部には、より大きな眼状斑がある。
ボルボックスは、協調と分業への最初のステップの例となることができる。 ボルボックスのコロニーは、さまざまな機能を果たすために分離された細胞からなる多細胞型と見なすことができる。 その機能とは、
- 食物の生産に関わる植物性細胞、運動に関わる細胞、
- 娘コロニーを作る無性細胞、
- 有性細胞、である。 375>
- 性細胞:卵を産み、葯を産む
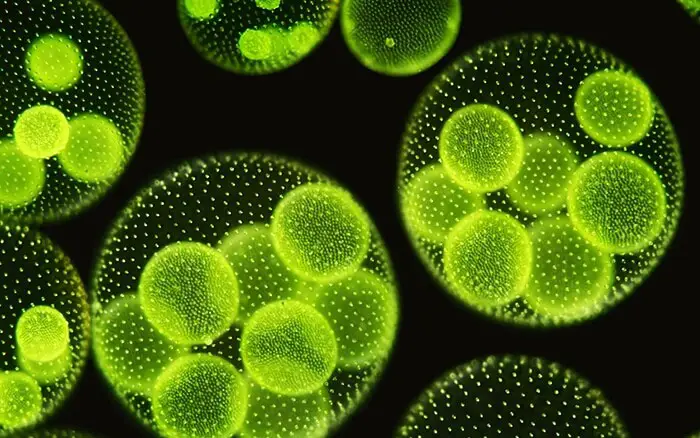
Volvox Under Microscope
Volvox Colony Under Microscope
Volvox Reproduction
Chlamydomonas とは異なり、ボルボックスのコロニーは細胞が機能分化していることが特徴です。 無性生殖と有性生殖の両方を行う。 生育期の初期(好条件)には無性生殖を行う。
Asexual Reproduction
Asexual reproductionは生育期の初期、好条件の時に起こる。 初期にはコロニーの細胞はすべて同じであるが,その後,コロニーの後半分の少数の細胞が餌を蓄えて大きくなる。 この大きく肥大した細胞は、淋菌(単数形淋菌)と呼ばれる特殊な無性細胞である。 その数は1つのコエノビウムで2〜50個とさまざまである。
淋菌からの娘コエノビウムの発生
分裂前の淋菌はコロニーの内部に少し押し込まれており、その大きさ、丸い形、鞭毛と眼点がない、目立つ核、いくつかのピレノイド、緻密な粒状の細胞質によって植物細胞列として区別されることができる。
生殖腺原基の第1分裂は、coenobiumの前後面を縦断している。 第2分裂も縦長で、第1分裂と直角である。 こうしてできた娘細胞はそれぞれ再び縦に分裂し、8細胞板が形成される。 8枚の細胞板は、凹面が外側を向くように湾曲する。 この8細胞期はプラケア期と呼ばれる。 16細胞期になると、細胞は中空球の周囲に配列され、親ケノビウムの外側に向かって小さな開口部であるフィアロポアが形成される。 この段階では、すべての細胞は裸で互いに接触している。 その前端は球の中心を向いている。 侵食の進行に伴い、鞘孔は大きく拡大する。 後部の折り畳み(侵入)がフィアロポアを押し始めると、その周囲の縁は後方にカールし、徐々に滑り落ち、構造全体が反転する。 いくつものひだを持つようになった孔は、次第に閉じていく。 反転には3〜5時間かかる。 異常な例では、V. minorで報告されているように、反転が全く起こらないこともある。
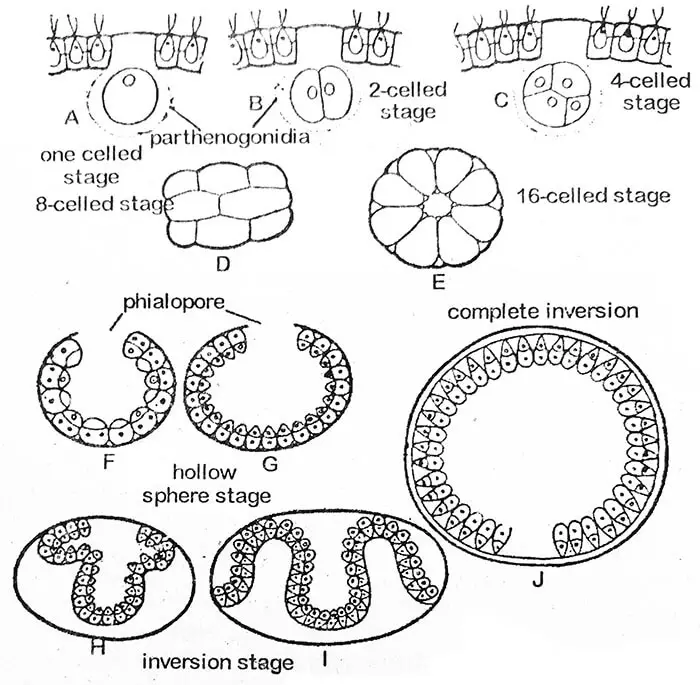
今度は娘コロニーの細胞が粘液性の部分(細胞壁)を発達させて互いに分離し始める。 そして、それぞれの細胞は、ついに一対の鞭毛と細胞膜を獲得する。 娘コロニー(コエノビウム)はまだ親細胞壁の中に留まっており、やがて粘液性の膜に包まれるように発達する。 1つの親コロニーに複数の娘コエノビウムが同時に発生することもある。
親コロニーの死滅と崩壊に伴い、娘コエノビウムは放出される。 時には娘コロニーが長く放たれずに孫コロニーを発達させることもある。
有性生殖
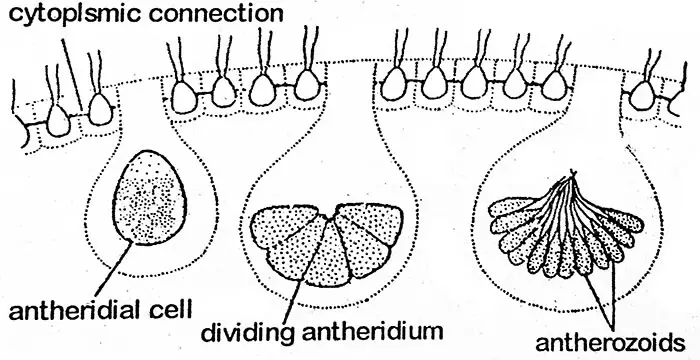
ボルボックスは高度な有性生殖を行い、葯とオゴニアが形成される。 これらはV. globatorのように同じコエノビウムに形成されるもの(単為生殖)とV. aureusのように異なるコエノビウムに形成されるもの(雌雄生殖)がある。 単子葉の種は原生種であるため、このような種では、異なる植物の葯と卵子の間で受精が起こる。 有性コロニーには、無性コロニーを形成しないことが多いのも興味深い。 コエビ類では、性器を形成する予定の細胞は、後半に存在する。 これは特殊な細胞であると考えられる。 性器(gametangia)の数は少ない。 配偶子が形成される過程で、細胞は肥大して丸みを帯び、鞭毛を捨てるが、他の細胞とは細い原形質の糸でつながったままである。
無性生殖細胞が娘コロニーになるのと同じように、子房の原形質が細胞分裂を繰り返す(plakea stage)。 このようにして、16〜512個の小さな裸の二鞭毛虫状の褐虫藻が葯の中に大量に形成される。 黄色ブドウ球菌のように無性コロニーで見られる場合を除き、これらは平板状に集合している。
アンテロゾイド
各アンテロゾイドは二鞭毛で細長い円錐形または牙状構造で、単一の核と小さな黄緑または淡緑の葉緑体を持っています
Oogonium
反すう細胞はかなり大きくなって鞭毛や隣接細胞との原形質の結合を捨てる。 細胞は丸みを帯びたり、フラスコ状になり、その大部分がコエノビウムの内部に突出する。 この段階で、細胞全体が片側に向かってくちばしのような突起を持つ単一の球形の卵に変化し、これをウゴニウムと呼ぶ。 この端からantherozoidがoogoniumの中に入り込む。 卵巣には、中央に大きな核と、ピレノイドを持つ葉緑体がある。 原形質を通じて隣接する細胞から吸収された予備物質がしばしば豊富に蓄えられている。
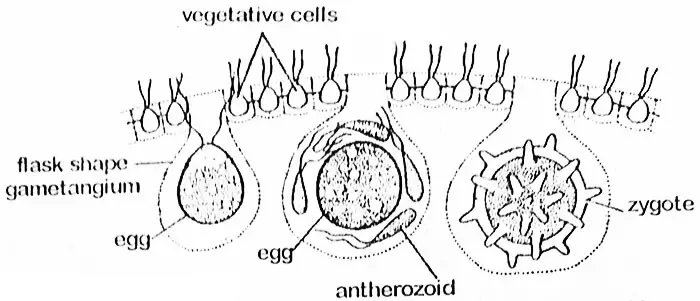
受精
受精すると、反すうは集団で放出されて、これが卵に到達するまでそのままの状態を維持する。 その後、antherozoidsは解放されます。 卵と融合し、胞子を形成するのは1個だけです。 卵胞はその後、3層構造の平滑または棘のある壁を分泌する。 ヘマトクロム(キサントフィル系の赤色色素顆粒)を十分に蓄積し、オレンジ色の外観を呈する。
胞子とその発芽
外胞子は平滑壁(V. globator)と棘状壁(V. speematospaera)がある。 中層は中胞子、内層は内胞子である。 接合体には十分な予備餌料とその他の包有物が含まれている。 接合体はコエノビウムによって保持され、ゼラチン質のマトリックスが崩壊することによって遊離することができる。 接合子は遊離後、プールの底に沈殿し、数年間は生存可能である。 A:発芽した胞子、B:胞子から解放されたスワーマー、C:スワーマーの最初の分裂、D:新しいコロニーの形成、E:侵入したコロニー、F:脱出可能なコロニー(図2.27)
好条件が発生すると接合体は様々に発達する。 V. campensisでは接合子の核は減数分裂して4個の核を形成し、そのうち3個は退化し、1個が生き残る。 生き残った核は、細胞質とともに小胞から脱出する。 この段階で、それはswarmer(昆虫、細胞などの多数または密集したグループ)と呼ぶことができ、自由に泳ぎ、分裂と再分裂を繰り返し、新しいcoenobiumを形成する。 発芽の際、外側の2つの壁層はゼラチン状になり、内側の層は小胞を形成し、後に接合体プロトプラストで満たされる。 V. rouselettiとV. minorでは、接合子のプロトプラストは単一の胞子に変換され、さらに分裂して新しいコエノビウムを形成する。 このようなコエノビウムは少数の細胞からなり、次の6世代以上にわたって無性生殖を行い、その都度、後継世代で数を増やしていく。 図2.28)
接合体はボルボックスのライフサイクルの中で唯一の二倍体であり、したがって植物本体はハプロイドである。 そのため、接合体は新しいコロニーを形成する際に還元分裂を行わなければならない
。