Gameten en bevruchting overzicht
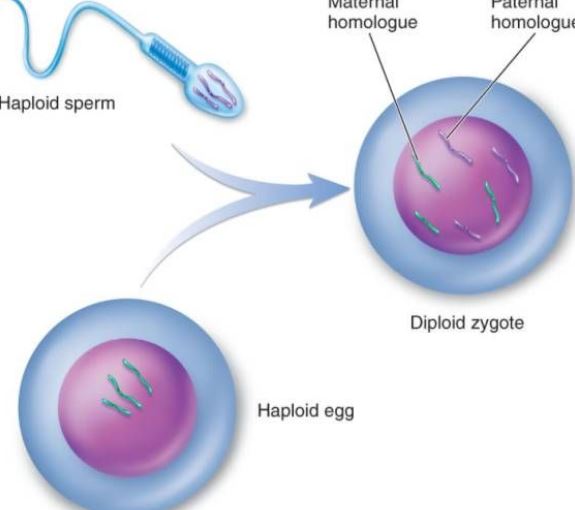
Seksuele voortplanting verwijst naar de seksuele geslachtscellen (gameten) die door de ouders worden geproduceerd, door de combinatie van biseksuele geslachtscellen (zoals sperma- en eicellen), worden de bevruchte eicel, en vervolgens ontwikkelt de bevruchte eicel zich tot een nieuw individu, de zogenaamde geslachtelijke voortplanting. Uitgebreide variatie in gencombinaties bij geslachtelijke voortplanting kan het vermogen van de nakomelingen om zich aan natuurlijke selectie aan te passen, vergroten. Willekeurig gecombineerde genen in nakomelingen van geslachtelijke voortplanting kunnen al dan niet gunstig zijn voor de soort, maar vergroten in ieder geval de kans dat enkele individuen overleven in een onvoorspelbaar en steeds veranderend milieu, wat de soort ten goede komt. Seksuele voortplanting kan ook de verspreiding van gunstige mutaties in de populatie bevorderen. Indien twee individuen in een soort gunstige mutaties hebben op verschillende plaatsen, zullen in de aseksuele voortplantingspopulatie de twee mutanten met elkaar wedijveren tot een eliminatie, het is onmogelijk om beide gunstige mutaties tegelijkertijd te behouden. In populaties van seksuele voortplanting echter kunnen deze twee gunstige mutaties door paring en recombinatie gelijktijdig in het genoom van hetzelfde individu terechtkomen en zich gelijktijdig in de populatie verspreiden. Om bovengenoemde redenen versnelt seksuele voortplanting het evolutieproces. In de meer dan 3 miljard jaar van biologische evolutie op aarde zijn de eerste twee miljard jaar van het leven in de fase van de ongeslachtelijke voortplanting gebleven en is de evolutie traag geweest; sinds de laatste 1 miljard jaar is het tempo van de evolutie aanzienlijk versneld. Naast veranderingen in het milieu op aarde (zoals het ontstaan van de zuurstofhoudende atmosfeer, enz.), is ook het ontstaan en de ontwikkeling van de geslachtelijke voortplanting een belangrijke oorzaak.
Gameten
Gameten verwijzen naar rijpe cellen die door het voortplantingssysteem worden geproduceerd wanneer het organisme zich geslachtelijk voortplant, kiemcellen genoemd. Gameten worden onderverdeeld in mannelijke gameten en vrouwelijke gameten. De vrouwelijke geslachtscellen van dieren en planten worden gewoonlijk eicellen genoemd, en de mannelijke geslachtscellen sperma. Sperma is vrij klein, maar het kan bewegen en in een schorpioenvorm de eicel binnendringen. De eicel is vrij groot en niet zwemmend. Het volume van de eicel van de zee-egel is bijvoorbeeld 10.000 maal groter dan dat van de zaadcellen. Hoewel de mannelijke en vrouwelijke geslachtscellen verschillend zijn in volume, is het kern-DNA dat zij leveren voor de nakomelingen gelijk, dat wil zeggen dat elk een set genomen levert. Door de grote omvang van de eicellen worden de cytoplasmatische structuur en het cytoplasmatische DNA van de dochtercellen echter hoofdzakelijk geleverd door de eicellen. Gameten spelen een belangrijke rol in biologische berekeningen. Via genetische kaarten kunnen de genenstroom en het patroon van de genotypen van de nakomelingen duidelijk worden waargenomen. Het aantal gameten in de eerste generatie hybriden en de relatieve eigenschappen in hybriden is de 2nde macht (n is de logaritme van de relatieve eigenschappen). Wanneer een levend lichaam een gameet vormt, worden de gepaarde genetische factoren van elkaar gescheiden en komen ze in verschillende gameten terecht. De gameten bevatten slechts één van elk paar genetische factoren. Bij de bevruchting is de combinatie van mannelijke en vrouwelijke gameten willekeurig. Als gevolg van het geslacht der gameten gevormd door meiose, de verscheidenheid van chromosoomsamenstelling, het verschil in genetisch materiaal van verschillende gameten, en de willekeur van de combinatie van eicellen en zaadcellen in het bevruchtingsproces, moeten de nakomelingen van dezelfde ouder divers zijn.
Fertilisatie
Fertilisatie is het proces waarbij eicellen en zaadcellen samensmelten tot één zygote. Het is een basiskenmerk van de geslachtelijke voortplanting en is alomtegenwoordig in de dieren- en plantenwereld, maar het wordt vaak aangeduid als het meest Op cellulair niveau omvat het bevruchtingsproces drie belangrijke stadia: activering van de eicel, regulering, en amfotere pronucleaire fusie. Activering kan worden beschouwd als het beginpunt van de ontogenese, die zich vooral manifesteert door veranderingen in de doorlaatbaarheid van de eicelmembraan, de ecclesie van de corticale granula, de vorming van het bevruchte membraan, enz. Aanpassing vindt plaats na activering, die de eerste stap is in het verzekeren van de normale deling van bevruchte eicellen. De verandering van de prokaryote kern verzekert de overerving van de ouders en herstelt de diploïd. De bevruchting brengt niet alleen de replicatie van het DNA op gang, maar activeert ook de genetische informatie zoals mRNA en rRNA in de eicel om de proteïnen te synthetiseren die nodig zijn voor de ontwikkeling van het embryo. Na 6 tot 7 dagen na de bevruchting verdwijnt de blastocyste zona pellucida en wordt deze geleidelijk begraven en bedekt door het baarmoederslijmvlies, wat innesteling van de bevruchte eicel wordt genoemd. Het sperma van een dier heeft geen duidelijke chemotaxis zoals het sperma van lagere planten zoals bryophyten, maar bereikt de omgeving van de eicel door actieve beweging of door de ciliaire beweging van de genitale epitheelcellen. De capacitatie van het sperma: Het is bekend dat wanneer veel zaadcellen van zoogdieren het vrouwelijke voortplantingskanaal of de cumulus passeren, het vreemde eiwit dat de zaadcellen inkapselt, wordt verwijderd en de fysische en biologische eigenschappen van het plasmamembraan van de zaadcellen veranderen, waardoor de zaadcellen energie kunnen winnen en aan het bevruchtingsproces kunnen deelnemen. Wanneer de zaadcel van een zoogdier in contact komt met het eimembraan of de zona pellucida van de eicel, bindt hij zich specifiek aan een glycoproteïne op het eimembraan, waardoor de zaadcel wordt gestimuleerd energie te produceren. De acrosoomreactie helpt het sperma om het eimembraan verder te doorkruisen. Op het zee-egel-ei is de acrosoomreactie van het sperma een bepaalde polysaccharidesubstantie in de peri-membraanfilm. De meeste eieren hebben eimembranen aan de periferie, en de dikte van de verschillende eimembranen varieert. Het hoofdbestanddeel is mucine of mucopolysaccharide; slechts enkele zijn naakte eieren, zoals de eieren van een coelenteraat. Acrosoomreactie: Wanneer het sperma door het eimembraan gaat, vindt er een proces van hechting plaats. Het eerste is losjes gehecht, vrij van externe temperatuurinterferentie, en heeft geen specificiteit. Tijdens de hechtingsperiode wordt het protoplast-eiwit op het acrosomale membraan omgezet in acrosomeiwit, en het acrosomeiwit versnelt de spermacel om het eimembraan te doorkruisen; het is een sterke combinatie, kan door lage temperatuur worden gestoord, en heeft een specificiteit. Op het plasmamembraan van het sperma van de zee-egel is een eiwit geïsoleerd dat zich specifiek aan het ei-glycoproteïne bindt, een bindend eiwit genaamd, met een molecuulgewicht van ongeveer 30.000. De secundaire eicellen in het midden van de tweede rijpingsdeling tijdens de ovulatie komen, samen met de omliggende zona pellucida en de stralingskroon, snel via de buikholte in de ampulla van de eileider terecht als gevolg van het slingeren van de trilhaartjes van de epitheelcellen van de eileider en het samentrekken van de spierlaag. Indien de eicel het sperma niet ontmoet, begint deze gewoonlijk binnen 12-24 uur te degenereren. Zodra het sperma in contact is met de eicel, ondergaat de eicel zelf een reeks activeringsveranderingen. Bij zoogdiereieren wordt deze gekenmerkt door de corticale reactie, de ovariële membraanreactie en de zona pellucida-reactie, die de multi-bevruchting kunnen tegenhouden en de verdere ontwikkeling van de eicellen kunnen stimuleren. De corticale reactie treedt op op het moment van de versmelting van de zaadcellen. Vanaf het punt van samensmelting scheuren de corticale deeltjes en wordt de inhoud uitgestoten, waardoor de cortex van de hele eicel wordt verspreid. De zona pellucida-reactie is een recombinatieproces van het omhulsel van het ovum en de corticale korrels. De zona pellucida-reactie is een proces waarbij de corticale uitvloeiing en de zona pellucida van een bevruchte eicel, de eicelmembraan wordt gescheiden van de plasmamembraan, de sperma-receptor in de zona pellucida verdwijnt, en de zona pellucida verhardt. Alleen sperma dat een acrosinereactie vertoont, kan met de eicel versmelten. Onder invloed van acrosine passeert het sperma de radiale kroon en treedt het in wisselwerking met de sperma-receptorglycoproteïnemolecule ZP3 op de zona pellucida, waardoor het sperma het acrosomale enzym kan loslaten en via de zona pellucida in de perivitelline terechtkomt. Bij het begin van de bevruchting komt het omhulsel van het equatoriale deel van de kop van de menselijke zaadcel in contact met het membraan van de eicel, waarna de kern en het cytoplasma van de zaadcel de eicel binnendringen. Nadat het sperma de eicel is binnengedrongen, laten de corticale deeltjes in het ondiepe cytoplasma van de eicel onmiddellijk hun inhoud los in de periferie van het membraan. Tegelijkertijd, wanneer het membraan van de corticale korrels versmelt met de eicellen, neemt de negatieve oppervlaktelading op het celoppervlak toe, waardoor de versmelting van het plasmamembraan van het sperma met het eimembraan wordt onderdrukt, hetgeen een corticale reactie wordt genoemd. De structuur van de zona pellucida verandert, wat de zona pellucida-reactie wordt genoemd. Op dat moment wordt het vermogen van de zona pellucida om sperma te binden verminderd, waardoor het optreden van polyspermie wordt voorkomen en de biologische kenmerken van menselijke monospermie worden gewaarborgd. Naast het vernietigen of inactiveren van de aan het sperma in de zona pellucida gebonden receptor, kan de inhoud van de corticale korrels ook de cross-linking tussen de peptideketens in de zona pellucida vergroten, de gevoeligheid voor het acrosoomprotease verzwakken en de penetratie van het sperma verhinderen. Hoewel er verschillende spermacellen door de zona pellucida gaan, dringt slechts één spermacel de eicel binnen om deze te bevruchten. In een abnormale situatie zijn er twee spermacellen bij de bevruchting betrokken, namelijk een dubbele spermabevruchting. Twee spermacellen gaan tegelijkertijd de eicel binnen en vormen embryo’s van triploïde cellen, die allemaal worden geaborteerd of kort na de geboorte sterven. Nadat het sperma de eicel is binnengedrongen, voltooit de eicel snel de tweede rijpe deling. Op dat moment worden de kernen van het sperma en de eicel respectievelijk de mannelijke pronucleus en de vrouwelijke pronucleus genoemd. De twee pronuclei naderen elkaar geleidelijk, het kernmembraan verdwijnt, en de chromosomen versmelten tot een diploïde bevruchte eicel. Wanneer de zaadcellen versmolten zijn, kan men zien dat de microvilli op het oppervlak van de eicel de zaadcellen omringen, die georiënteerd kunnen zijn; dan versmelt het eierstokmembraan met het plasmamembraan in het achterste gebied van het acrosoom van de zaadcellen.
Referentie
- Yanagimachi R. Kiemcellen en bevruchting: waarom ik deze onderwerpen bestudeerde en wat ik leerde op de weg van mijn studie. Andrologie. 2014, 2(6):787-93.
- Henshaw J M, Marshall D J, Jennions M D, et al. Lokale gametencompetitie verklaart geslachtstoewijzing en bevruchtingsstrategieën in de zee. American Naturalist. 2014, 184(2):32.
- Mori T, Igawa T. Gamete attachment process revealed in flowering plant fertilization. Plant Signaling & Behavior. 2014, 9(12):977715.
- Hédouin L, Pilon R, Puisay A. Hyposalinity stress compromises the fertilization of gametes more than the survival of coral larvae. Marine Environmental Research. 2015, 104:1-9.
Luo J, Mcginnis L K, Carlton C, et al. PTK2b function during fertilization of the mouse oocyte. Biochem Biophys Res Commun. 2014, 450(3):1212-1217.