ADVERTENTIES:
In dit artikel gaan we in op Paramecium Caudatum:- 1. Habitat, habitat en cultuur van Paramecium Caudatum 2. Structuur van Paramecium Caudatum 3. Structuur van Paramecium Caudatum 3. Voortbeweging 4. Voeding 5. 5. Ademhaling en uitscheiding 6. Osmoregulatie Osmoregulatie 7. Gedrag 8. Voortplanting 9. Afwijkend gedrag bij de voortplanting 10. Enkele Cytoplasmatische Deeltjes.
Inhoud:
- Habitat, Habitat en Cultuur van Paramecium Caudatum
- Structuur van Paramecium Caudatum
- Locomotie van Paramecium Caudatum
- Nutritie bij Paramecium Caudatum
- Respiratie en Excretie bij Paramecium Caudatum
- Osmoregulatie van Paramecium Caudatum
- Gedrag van Paramecium Caudatum
- Reproductie bij Paramecium Caudatum
- Aberrant gedrag bij de voortplanting bij Paramecium Caudatum
- Enkele Cytoplasmatische Deeltjes Gemeld bij Paramecium Caudatum
- Habitat, Habitat en Cultuur van Paramecium Caudatum:
- Kweek van Paramecium:
- Structuur van Paramecium Caudatum:
- (i) Grootte en vorm van Paramecium Caudatum:
- (ii) Pellicula:
- (iii) Cilia:
- (iv) Infra-Ciliair Systeem:
- (v) Mondgroef en Cytopyge:
- (vi) Cytoplasma:
- (vii) Ectoplasma:
- (viii) Trichocysten:
- (ix) Neuromotorium en geassocieerde fibrillen:
- (x) Endoplasma:
- (xi) Contractiele vacuolen:
- (xii) Voedsel vacuolen:
- Locomotie van Paramecium Caudatum:
- (i) Metabolie of lichaamscontorties:
- (ii) Ciliaire Locomotie:
- Voeding van Paramecium Caudatum:
- Voedingsmechanisme:
- Vertering en Egestie van Paramecium Caudatum:
- Respiratie en Excretie van Paramecium Caudatum:
- Gedrag van Paramecium Caudatum:
- (i) Reacties op contact (Thigmotaxis):
- (ii) Reacties op chemicaliën (Chemotaxis):
- (iii) Reacties op temperatuur (Thermotaxis):
- (iv) Reacties op licht (Phototaxis):
- (v) Reacties op elektrische stroom (Galvanotaxis):
- (vi) Reacties op waterstroom (Rheotaxis):
- (vii) Reacties op de zwaartekracht (Geotaxis):
- Hervoortplanting bij Paramecium Caudatam:
- (i) Transverse Binaire Splijting:
- (ii) Conjugatie:
- Aberrant gedrag bij de voortplanting in Paramecium Caudatum:
- (i) Endomixis:
- (ii) Autogamie:
- (iii) Hemixis:
- (iv) Cytogamie:
- Enkele Cytoplasmatische Deeltjes Gemeld in Paramecium Caudatum:
- (i) Kappa Deeltjes:
- (ii) mµ, Deeltjes:
- (iii) Pi-deeltjes:
- (iv) Lambda Deeltjes:
Habitat, Habitat en Cultuur van Paramecium Caudatum:
Paramecium caudatum (Gr., paramekes = langwerpig; L., caudata = staart) wordt algemeen aangetroffen in zoetwatervijvers, poelen, sloten, beken, meren, stuwmeren en rivieren. Hij wordt vooral veel aangetroffen in stilstaande vijvers die rijk zijn aan rottend materiaal, in organische infusies, en in rioolwater. Paramecium caudatum is een vrij levend organisme en deze soort is wereldwijd verspreid.
Kweek van Paramecium:
ADVERTENTIES:
Neem ondergedompeld onkruid uit een vijver en doe dit in een pot met gedestilleerd water, dek de pot af en laat het staan om te rotten; zwermen Paramecia zullen in een paar dagen verschijnen. Kook nu hooi in water, giet het aftreksel af en voeg een paar tarwekorrels toe, en laat het staan tot het troebel is van de bacteriën.
Schenk Paramecia uit het eerste potje over in deze vloeistof, waar ze zich snel zullen vermenigvuldigen. Hooi infusies alleen zal produceren Paramecia met de aanwezigheid van cysten, en cysten die lijken op zandkorrels zijn gemeld, maar er is geen bewijs van Paramecia vormen cysten, omdat ze nooit zijn bevestigd.
Structuur van Paramecium Caudatum:
(i) Grootte en vorm van Paramecium Caudatum:
Paramecium caudatum (Fig. 20.1) is een microscopisch organisme en met het blote oog zichtbaar als een minuscuul langwerpig lichaam.
ADVERTENTIES:
Het ziet er lichtgrijs of wit uit en meet gewoonlijk tussen 170 en 290 micron in lengte en kan een lengte tot 300-350 micron bereiken. P. caudatum lijkt op de zool van een pantoffel of schoen, vandaar dat het dier algemeen bekend staat als pantoffeldiertje. Het is vier maal zo lang als breed en enigszins cilindrisch met duidelijk verschillende uiteinden.
Het voorwaarts bewegende voorste deel is slank met een stomp of afgerond uiteinde, terwijl het achterste uiteinde enigszins spits of kegelvormig is. Het breedste deel van het organisme bevindt zich juist onder het midden. Het lichaam van het dier is asymmetrisch van vorm en vertoont een goed afgetekend mond- of buikoppervlak en een aboraal of dorsaal oppervlak.
(ii) Pellicula:
Het lichaam is bedekt met een dunne, dubbelgelaagde, elastische en stevige pellicula, gemaakt van gelatine. De pellicula houdt de vorm van het dier, maar is elastisch genoeg om samentrekkingen mogelijk te maken. De pellicula heeft een dubbel membraan, waarvan het buitenste membraan doorloopt met de trilharen en het binnenste membraan met het ectoplasma. Onder de sterkere vergroting van de microscoop vertoont de pellicula rechthoekige of zeshoekige depressies op zijn oppervlak.

Deze rangschikking is bewaard gebleven op het dorsale oppervlak van Paramecium, maar op het ventrale oppervlak komen de ribbels voor en achter samen in de richting van een preorale en postorale apertuur. Elke zeshoekige depressie wordt geperforeerd door een centrale opening waardoor een enkel cilium naar buiten komt. De voorste en achterste marges van de zeshoekige depressies dragen de openingen van trichocysten.
Uit de elektronenmicroscopische studie van pellicula (Fig. 20.2) door Ehret en Powers (1957) is gebleken dat de zeshoekige depressies overeenkomen met regelmatige reeksen holten, de alveoli. Alle alveoli tezamen vormen een ononderbroken alveolaire laag, die wordt begrensd door een buitenste alveolaire en binnenste alveolaire membranen.
De buitenste laag ligt in nauw contact onder het buitenste celmembraan. Daarom omvat pellicle buitencelmembraan, buitenste alveolaire membraan en binnenste alveolaire membraan.
(iii) Cilia:
Het gehele lichaam is bedekt met talrijke, kleine, haarachtige uitsteeksels, cilia genaamd. Cilia komen voor in longitudinale rijen over het hele lichaam, deze toestand staat bekend als holotrichous waarbij de lichaam cilia gelijk zijn. Cilia hebben dezelfde structuur als flagellen, ze hebben een buitenste protoplasmatische schede of plasmamembraan met negen dubbele overlangse fibrillen in een perifere ring. Bij sommige cilia zijn de negen buitenste fibrillen niet gepaard.
ADVERTENTIES:
Er zijn twee centrale longitudinale fibrillen die dunner zijn dan de buitenste fibrillen. Elk cilium komt voort uit een basale korrel of kinetosoom. De negen paren perifere fibrillen smelten samen om de wand van het kinetosoom te vormen, zodat het kinetosoom een buis is die aan het onderste uiteinde open of gesloten is; bij de meeste ciliaten houden de twee centrale fibrillen op ter hoogte van de pellikel.
Uit het kinetosoom komt een dunne rhizoplast die zich niet met de kern verbindt. Veel Metazoa hebben ook cilia, hun structuur is hetzelfde, behalve dat de basale korrel anders is en fijne filamenten of wortelvezels heeft die tot in het cytoplasma reiken. Maar cilia verschillen van flagella doordat ze over het algemeen talrijker zijn en korter van omvang.
De ciliatuur kan gemakshalve worden verdeeld in lichaams- of somatische ciliën, die zich op het lichaamsoppervlak bevinden, en in orale ciliën, die met het mondgebied zijn verbonden. De lichaamshaarcellen zijn gelijk, maar zijn langer aan het achterste uiteinde, vandaar de naam caudatum. De trilharen zijn organellen voor de voortbeweging en het verzamelen van voedsel, en fungeren ook als zintuigreceptoren en detecteren de prikkels van de externe omgeving.

Ultrastructuur van Cilia:
De cilia en flagella hebben een fibrillaire samenstelling. Aan de basis heeft het cilium een diameter van ongeveer 0,2 micron of 2.000 A0 die tot 10 micron boven het celoppervlak kan liggen. De cilia zijn begrensd door een eenheidsmembraan van 90 A0 dikte dat lijkt op en continu blijft met het plasmamembraan. De begrensde ruimte van het cilium bevat een waterige substantie die matrix wordt genoemd.
In de matrix blijven elf longitudinale fibrillen of microtubuli ingebed. Van de elf fibrillen bevinden zich er twee in het centrum, terwijl de overige negen fibrillen perifeer rond de centrale fibrillen gerangschikt blijven. Elk van de negen buitenste fibrillen heeft een diameter van 360 A0 en is samengesteld uit twee sub-fibrillen met een diameter van 180 tot 250 A0.
Deze sub-fibrillen worden aangeduid als de sub-fibril A en sub-fibril B.
ADVERTENTIES:
De sub-fibril A is iets groter dan de sub-fibril B. De sub-fibril A geeft aan zijn ene zijde twee dikke uitsteeksels of armen. De armen van de sub-fibril A van alle buitenste fibrillen blijven in de richting van de wijzers van de klok gericht. Verder ligt de sub-fibril A dichter bij het centrum van het cilium dan de sub-fibril B. Beide sub-fibrillen hebben een gemeenschappelijke wand van 50A0 dikte.
De twee centrale fibrillen hebben geen gepaarde sub-fibrillen zoals de perifere negen fibrillen, maar bevatten elk slechts een enkele tubulus. Elke centrale fibril heeft een diameter van ongeveer 250A0 en is samengesteld uit 60A0 dikke wand.
De beide centrale fibrillen blijven gescheiden door een ruimte van 350A0 en blijven gehuld in een gemeenschappelijke schede. Gibbnos (1967) heeft gerapporteerd dat de schede van de centrale fibrillen negen radiaal georiënteerde schakels of spaken aan elke sub-fibril A afgeeft.

De hoge resolutie elektronenmicroscopie heeft aangetoond dat elk van de perifere en centrale fibrillen van de cilia en flagella is samengesteld uit tien tot twaalf filamenten van 40A0 dikte. Elk filament is gepareld. Elke parel blijft gerangschikt in rasters van 40 bij 50A0 in het vlak van de wand van de tubulus. Deze kralen worden beschouwd als de basis subeenheid van de tubule structuur.
(iv) Infra-Ciliair Systeem:
Het infra-ciliair systeem bevindt zich vlak onder de pelliculaire alveoli. Het bestaat uit het kinetosoom of basaal lichaam en kinetodesma. De cilia ontstaan uit kinetosomen en uit elk kinetosoom ontstaat een delicaat cytoplasmatisch bindweefsel dat kinetodesma wordt genoemd, (Fig. 20.2).
Ligt onder de pellicula iets naar rechts, maar verbonden met alle kinetosomen van één longitudinale rij, is een longitudinale bundel van verscheidene kinetodesmata, een kinetodesmata van elk kinetosoom strekt zich over een afstand anterieur uit in zijn eigen bundel van kinetodesmata.
Een longitudinale rij kinetosomen met hun kinetodesmata vormt een longitudinale eenheid die een kinetie wordt genoemd. Alle kineties of kinetia vormen het infra-ciliaire systeem van een ciliate. De kinetia liggen in de cortex onder de pellicula, hun aantal is vrij constant voor elke ciliate.
Het infra-ciliair systeem controleert en coördineert de bewegingen van de cilia, en het zorgt voor de vorming van organellen bij celdeling, b.v. sommige kinetia vormen de mond. Bij de binaire splitsing van ciliaten worden de kinetia dwars doorgesneden in twee, die elk naar één dochtercel gaan, dit wordt perikinetale splitsing genoemd.
(v) Mondgroef en Cytopyge:
Aan de ventrolaterale zijde is een grote schuine, ondiepe depressie, mondgroef of peristoom genoemd, die het dier een asymmetrisch uiterlijk geeft. Hij loopt schuin van één kant naar achteren (meestal van links naar rechts, maar in sommige gevallen van rechts naar links) en eindigt iets achter het middenlichaam. De mondgroef mondt uit in een korte kegelvormige trechtervormige holte, vestibule genaamd.
ADVERTENTIES:
De vestibule mondt rechtstreeks uit in de vaste, ovaalvormige opening die cytostome (mond) wordt genoemd. Rechtstreeks vanuit het cytostoma strekt zich de brede cytopharynx uit naar het centrum van het lichaam. De cytopharynx draait vervolgens scherp naar de achterzijde om over te gaan in de slanke taps toelopende oesophagus.
Dus is de slokdarm ongeveer evenwijdig aan het lichaamsoppervlak van Paramecium, behalve aan het achterste uiteinde. Hier draait de slokdarm weer naar het centrum van het dier om in de zich vormende voedselvacuole uit te monden.
De cytopyge (ook wel celanus of anale vlek of cytoproct genoemd) ligt op het ventrale oppervlak van het lichaam bijna verticaal achter de cytostome of mond. Onverteerde voedseldeeltjes worden via de cytopyge uitgescheiden. De ciliatie van de cytopharynx is zeer gecompliceerd.
Gelei (1934) meldde de aanwezigheid van vier rijen en Lund (1941) nam ten minste vier rijen waar. Een structuur, penniculus genaamd, bevindt zich op de linkerwand van de cytofarynx en maakt een spiraal van ongeveer 90 graden, zodat het achterste uiteinde zich op het orale (ventrale) oppervlak van de slokdarm bevindt.
Volgens Lund bestaat de penniculus uit acht rijen cilia, gerangschikt in twee dicht op elkaar staande blokken van elk. Een soortgelijke band, bestaande uit vier rijen lange cilia die minder compact zijn dan bij de penniculus, wordt de quadrulus genoemd. Deze loopt spiraalsgewijs langs de dorsale wand van de mondholte en eindigt dicht bij de penniculus.
De penniculus en quadrulus zijn door sommige werkers ten onrechte een golvend membraan genoemd. De quadrulus en penniculus regelen de passage van voedsel. Het is niet bekend hoe cilia werken, waarschijnlijk trekken hun fibrillen zich op ritmische wijze samen waardoor een buiging ontstaat. Gelei (1925) wees erop dat de functie van de penniculus is om voedselelementen het lichaam in te duwen.

(vi) Cytoplasma:
Het cytoplasma is onderscheiden in een smalle, uitwendige of corticale zone die het ectoplasma wordt genoemd en een groter, inwendig of medullair gebied dat het endoplasma wordt genoemd.
(vii) Ectoplasma:
Het ectoplasma (ectosarc of cortex) is een blijvend deel van het lichaam, dat opvallend is afgescheiden van het endoplasma. Het ectoplasma vormt een stevige, heldere, dunne en dichte buitenlaag. Het bevat de trichocysten, cilia en fibrillaire structuren en is aan de buitenkant begrensd door een bedekking, pellikel genaamd.
(viii) Trichocysten:
Ingebed in het ectoplasma, loodrecht op het oppervlak, zijn kleine spoelvormige zakjes, trichocysten genaamd. Een kleine vlek op elke voorste en achterste rand van de zeshoek markeert de plaats van een trichocyst. Ze zijn gevuld met een lichtbrekende, dichte vloeistof met een zwellende substantie, aan het buitenste uiteinde zit een kegelvormige kop of stekel.
De trichocysten liggen loodrecht in het ectoplasma, ze openen zich door kleine poriën op de richels van de zeshoekige gebieden van de pellicula.
Ze ontstaan uit kinetosomen van cilia, migreren dan en vestigen zich op gelijke afstand in het endoplasma. Wanneer het dier geïrriteerd is, worden de trichocysten als lange kleverige draden uitgestoten. Een geloste trichocyt heeft een ondoorzichtige spike, zoals een omgekeerde spijker, en een lange gestreepte schacht, maar de schacht is niet te zien in niet-geloste toestand en wordt waarschijnlijk gevormd tijdens de ontlading.
De functie van trichocysten is onzeker, maar ze worden ontladen als reactie op plaatselijke contacten en verwondingen, ze kunnen dienen als organellen van verdediging.
Maar dit is onzeker omdat de trichocysten ineffectief zijn tegen Didinium, de voornaamste predator op Paramecium, ze kunnen dienen om het diertje aan een plek te fixeren tijdens het voeden. Bij sommige ciliaten fungeren de trichocysten als aanvalsorganellen. Nadat de trichocysten zijn geloosd, worden ze geregenereerd uit kinetosomen.


(ix) Neuromotorium en geassocieerde fibrillen:
Volgens Lund (1933) bevindt zich op de linker dorsale wand van de cytofarynx ongeveer ter hoogte van de achterrand van het cytostoom een zeer kleine, tweebobbige massa, het neuromotorium. Vanuit het neuromotorium stralen fibrillen uit in het endoplasma.
Van deze gaan er gewoonlijk vier of meer tot bijna aan de dorsale lichaamswand, maar de rest is korter en niet definitief in positie. Alle worden endoplasmatische fibrillen genoemd. Hun functie is onbekend, maar mogelijk coördineren zij de voedingsbewegingen van de orale cilia. De fibrillen kunnen ook mechanische steun, elasticiteit, contractiliteit, geleiding en metabole invloed geven.

(x) Endoplasma:
Het endoplasma of medulla is het vloeibaarder en volumineuzer gedeelte van het cytoplasma, dat vele cytoplasmakorrels bevat, alsmede andere insluitsels en structuren van een gespecialiseerde aard. De cytoplasmatische insluitsels zijn mitochondriën, Golgi-apparaten, vacuole, kristallen, granules en chromidia, enz. Andere structuren, nl. kernen, samentrekkende vacuolen en voedselvacuolen worden eveneens in het endoplasma aangetroffen.
Nuclei:
In het endoplasma bij het cytostoom bevinden zich twee kernen, d.w.z., Paramecium is heterokaryotisch, een grote ellipsoïdale en korrelige macronucleus en andere kleine compacte micronucleus.
Macronucleus is een opvallend, ellipsoïdaal of niervormig lichaam. Het is van een compact type met fijne draden en dicht opeengepakte discrete chromatinekorrels van variabele grootte en ingebed in een achromatische matrix. Het bezit veel nucleoli en veel meer chromatinemateriaal (DNA).
Het is een somatische of vegetatieve kern. Hij deelt zich amitotisch en controleert de vegetatieve functies (metabolische activiteiten) van het dier. Hij ondergaat wel mitose.
Micronucleus is klein, compact en bolvormig. Hij bevindt zich meestal dicht bij de macronucleus, vaak in een concaviteit. Fijne chromatinekorrels en -draden zijn gelijkmatig over de structuur verdeeld. De micronucleus deelt zich mitotisch en controleert de voortplanting.
Het aantal micronucleus varieert met de soort; het is één bij P. caudatum, twee bij P. aurelia en vele bij P. multimicronucleatum. De micronucleus bevat een duidelijke nucleolus bij P. aurelia, maar deze wordt niet aangetroffen bij P. caudatum. Moses (1949; 1950) heeft gemeld dat de macronucleus en de micronucleus identiek zijn in chemische samenstelling.
(xi) Contractiele vacuolen:
Er zijn twee grote, met vloeistof gevulde contractiele vacuolen, elk gelegen bij een uiteinde van het lichaam dicht bij het dorsale oppervlak. Hun positie is vast (in tegenstelling tot Amoeba), zij liggen tussen het ectoplasma en het endoplasma, maar het zijn tijdelijke organellen die periodiek verdwijnen. Bij sommige soorten schijnen zij een bekledingsmembraan te hebben, in welk geval zij niet geheel verdwijnen tijdens de systole.
Verbonden met elke samentrekkende vacuole zijn vijf tot twaalf buisvormige uitstralende kanalen, elk bestaande uit een eindgedeelte, een lange ampulla die in elkaar zakt als hij leeg is, en een kort injectorkanaal dat uitkomt in de vacuole.

De kanalen staan in verbinding met een groot deel van het lichaam van waaruit zij vloeistoffen opnemen en in de vacuole gieten die aldus opnieuw wordt samengesteld en in omvang toeneemt, wanneer de samentrekkende vacuole zijn maximale omvang bereikt, trekt hij plotseling samen (systole) en loost zijn inhoud door een permanente porie in de pellikel, dan vormen de kanalen opnieuw de samentrekkende vacuolen, de kanalen verdwijnen niet geheel aangezien zij permanente structuren zijn.
De twee samentrekkende vacuolen ontladen zich onregelmatig, de achterste trekt sneller samen omdat deze zich dicht bij de cytofarynx bevindt en er meer water in komt. De hoofdfunctie van de kanalen en de samentrekkende vacuolen is hydrostatisch, zij verwijderen een teveel aan water uit het protoplasma, het water wordt gedeeltelijk geabsorbeerd en gedeeltelijk opgenomen tijdens de voeding.
De stikstofhoudende afvalstoffen bestaan uit ammoniakverbindingen en wat uraat, die samen met CO2 uit de samentrekkende vacuolen worden uitgestoten, maar er zijn geen aanwijzingen dat de uitscheidingsmaterie door het protoplasma in de kanalen of de samentrekkende vacuolen wordt uitgescheiden.
Er is een vaste permanente cytopyge of anus onder de peristome aan één kant van de cytopharynx, waardoor onverteerde voedselresten en uraten worden uitgescheiden.
De elektronenmicroscopische studie van de contractiele vacuolen heeft uitgewezen dat elk contractiel apparaat bestaat uit een deel van de tubuli van het endoplasmatisch reticulum, nefridiale tubuli, voedingskanalen, accessoire vacuolen (radiale kanalen) en de belangrijkste contractiele vacuole. De accessoire vacuolen worden verondersteld de ampullae van de voederkanalen te zijn (Fig. 20.11).
(xii) Voedsel vacuolen:
Dit zijn ruwweg bolvormige, niet-contractiele lichamen, variërend in grootte en aantal, die in het endoplasma liggen. Zij bevatten ingeslikte voedseldeeltjes, hoofdzakelijk bacteriën en een kleine hoeveelheid vloeistof, begrensd door een dun, definitief membraan. Volkonsky (1934) stelde de naam gastriolen voor deze vacuolen voor. Geassocieerd met de voedsel vacuolen zijn de spijsverteringskorrels.
Locomotie van Paramecium Caudatum:
Paramecium Caudatum beweegt zich voort op twee manieren, nl. door metabolische of lichaamsverdraaiingen en door cilia.
(i) Metabolie of lichaamscontorties:
Het lichaam van Paramecium Caudatum bezit elasticiteit, het kan zich door een doorgang wringen die nauwer is dan zijn lichaam, waarna het lichaam zijn normale vorm weer aanneemt. Deze tijdelijke verandering van lichaamsvorm is metabolisme, zij wordt in Paramecium teweeggebracht door het protoplasma.
(ii) Ciliaire Locomotie:
Locomotie bewerkstelligd door cilia is de voornaamste methode. De trilharen kunnen naar voren of naar achteren slaan, waardoor het dier naar voren of naar achteren kan zwemmen.
Normaal zwemt het dier voorwaarts, de trilhaartjes slaan schuin naar achteren, de trilhaartjes verstijven en buigen snel naar achteren tot ze bijna het lichaamsoppervlak raken, dit wordt de effectieve slag genoemd; daarna worden de trilhaartjes slap en keren langzaam terug naar de oorspronkelijke verticale positie, dit wordt de herstelslag genoemd.
Cilia van dezelfde transversale rij slaan samen en die van dezelfde longitudinale rij slaan na elkaar van het voorste naar het achterste uiteinde.

Deze gecoördineerde beweging van cilia wordt metachronaal ritme genoemd, dat te danken is aan het infra-ciliair systeem; dit veroorzaakt het voorwaarts zwemmen van het dier. Maar wanneer de lichaamsschelpen schuin naar achteren slaan, dan slaan tegelijkertijd de langere cilia van de mondgroef krachtiger, waardoor het voorste uiteinde naar links uitwijkt.
De werking van trilhaartjes van lichaam en mondgroef doet het dier om zijn lange as roteren. Deze draaiing is altijd naar links (behalve bij P. calkinsi die in een rechtse spiraal draait).
Deze combinatie van voorwaartse beweging, zwenken en rotatie doet het dier voorwaarts bewegen in een tegen de klok in draaiende spiraalbaan. Dit pad heeft een rechte as, en hetzelfde lichaamsoppervlak van het dier blijft in de richting van de as van het spiraalvormige pad. Maar bij het achteruit zwemmen, draaien alle soorten naar rechts.
De ciliaire slag kan worden omgekeerd, zodat de cilia schuin naar voren bewegen waardoor het dier achteruit zwemt. Door de ciliaire werking beweegt Paramecium met een snelheid van 1500 micron of zelfs meer per seconde.
Jennings beweert dat de spiraalvormige beweging van Paramecium te wijten is aan het feit dat de trilhaartjes voornamelijk naar achteren slaan, maar dat zij dit schuin naar rechts doen, waardoor het diertje naar links omrolt.
Ook dit zwenken van het lichaam naar het aborale oppervlak is grotendeels te wijten aan de grotere kracht van de effectieve slag van de orale trilhaartjes die directer naar achteren slaan. Het resultaat – de draaiing van het Paramecium om zijn lange as – stelt het Paramecium in staat een min of meer rechte koers te volgen bij het vormen van grote spiralen.
Voeding van Paramecium Caudatum:
In Paramecium Caudatum, is de voeding holozoïsch. Het voedsel bestaat voornamelijk uit bacteriën en minuscule Protozoa. Paramecium wacht niet op het voedsel, maar jaagt er actief op.
Er wordt beweerd dat Paramecium Caudatum een keuze in de keuze van zijn voedsel laat zien, maar daar lijkt geen basis voor te zijn, hoewel hij slechts bepaalde soorten bacteriën opslokt; de beschikbare gegevens suggereren dat 2 tot 5 miljoen individuen van Bacillus coli door een enkele Paramecium in 24 uur worden verslonden. Hij voedt zich ook met eencellige planten zoals algen, diatomeeën, enz. en kleine stukjes van dieren en groenten.
Voedingsmechanisme:
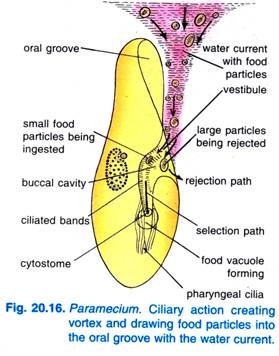
Wanneer Paramecium Caudatum in een gebied komt waar overvloedig voedsel is, komt hij tot rust. Hij voedt zich alleen in rust of wanneer hij heel langzaam zwemt, hij voedt zich nooit wanneer hij snel zwemt. Door het kloppen van de trilhaartjes van de mondgroef wordt een kegelvormige draaikolk van voedselrijk water in de mondgroef geslingerd op een afstand vóór het voorste uiteinde (Fig. 20.16).
De voedseldeeltjes gaan vervolgens naar de vestibule, van waaruit sommige voedseldeeltjes worden afgestoten en weggeworpen, maar andere in het cytostoom terechtkomen.
Aan het eind van de cytofarynx wordt een voedselvacuole gevormd die gevuld wordt met voedseldeeltjes. De quadrulus en peniculi regelen de passage van voedsel in de voedselvacuole die lateraal wordt gevormd. Wanneer de voedselvacuole een bepaalde grootte bereikt, omklemmen de post-buccale vezels de voedselvacuole en wordt deze door hen afgeknepen en op zijn beloop gelaten.
De vacuole bevat naast het voedsel ook wat water. Ronddraaiende bewegingen van endoplasma, cyclosis genoemd, voeren de voedselvacuole langs een bepaalde baan, die functioneel gelijkwaardig is aan een spijsverteringskanaal.
Het kanaal begint aan het einde van de cytofarynx, dan naar de achterzijde, dan naar voren om te circuleren met het endoplasma, dan naar het dorsale oppervlak, dan naar het voorste einde, dan naar beneden naar de cytopyge. In het begin van zijn reis neemt de voedselvacuole in omvang af, daarna neemt hij weer toe.

Vertering en Egestie van Paramecium Caudatum:
Tijdens cyclose vindt vertering plaats door enzymen die door het protoplasma in de vacuolen worden afgescheiden. Bij de vertering worden eiwitten omgezet in aminozuren, koolhydraten in oplosbare suikers en glycogeen, en waarschijnlijk worden ook vetten verteerd.
De inhoud van de voedselvacuolen is eerst zuur (pH ongeveer 4) en wordt daarna alkalisch; de belangrijkste vertering vindt plaats tijdens de alkalische fase. De onverteerde materie wordt met enige kracht door de cytopyge uitgescheiden.
Cyclose kan experimenteel worden aangetoond; als melk gekleurd met Congo rood wordt gevoerd aan Paramecium, zullen de vetbolletjes van melk in de voedsel vacuolen eerst rood worden door de zure reactie van enzymen, dan zullen ze veranderen van paarse tinten naar blauw door de alkalische reactie, de vacuolen zullen het verloop van cyclose laten zien.
Respiratie en Excretie van Paramecium Caudatum:
De uitwisseling van gassen (zuurstof en kooldioxide) vindt plaats door de semi-permeabele pellicula zoals bij andere zoetwaterprotozoën door het proces van diffusie. Paramecium Caudatum haalt zijn zuurstof uit het omringende water. Kooldioxide en organische afvalstoffen zoals ammoniak, afkomstig van de stofwisseling, worden waarschijnlijk uitgescheiden door diffusie naar buiten in het water in omgekeerde richting.

6. Osmoregulatie in Paramecium Caudatum:
Paramecium Caudatum heeft twee samentrekkende vacuolen, een anterieure en een posterieure. De functie van de samentrekkende vacuolen is osmoregulatie, d.w.z. het regelen van het watergehalte in het lichaam en kunnen ook dienen bij de uitscheiding van stikstofhoudende afvalstoffen zoals ureum en ammoniak.
Overschot aan water (door voortdurende endosmose) in het cytoplasma wordt afgescheiden in de buisjes van het endoplasmatisch reticulum en gaat naar de nefridiale buisjes → voedingskanalen → en verzamelt zich in de ampulla van een serie van 6 tot 11 uitstralende kanalen die convergeren naar en uitmonden in elke vacuole. De kanalen zijn het opvallendst als een vacuole wordt gevormd.
Wanneer elke vacuole is opgezwollen (diastole) tot een bepaalde grootte, trekt hij samen (systole) en loost naar buiten, waarschijnlijk via een porie. De samentrekkende vacuolen trekken om de beurt samen, met tussenpozen van 10 – 20 seconden.
De achterste contractiele vacuole werkt sneller dan de voorste vacuole vanwege de opname van grote hoeveelheden water in het achterste gebied door de cytofarynx. De contractiele vacuolen handhaven een optimale concentratie van water in het cytoplasma van het lichaam door het teveel af te voeren.

Gedrag van Paramecium Caudatum:
De reacties van Paramecium Caudatum op verschillende soorten prikkels worden geleerd door bestudering van zijn reacties en van de groepering of verstrooiing van individuen in een kweek. De reactie is positief als het dier naar een stimulus toe beweegt en negatief als het zich verwijdert. Op een ongunstige stimulus blijft het dier de vermijdende reactie geven tot het ontsnapt.
In de ontwijkende reactie keert de ciliaire slag om, beweegt het dier zich een korte afstand naar achteren, en roteert dan in een kegelvormige baan door het voorste uiteinde aboraal te zwenken terwijl het draait op het achterste uiteinde. Alle aanpassingen gebeuren met vallen en opstaan. Experimenten hebben aangetoond dat het voorste uiteinde van het dier gevoeliger is dan de andere delen.
De reacties van Paramecium op verschillende stimuli kunnen als volgt worden gegroepeerd:
(i) Reacties op contact (Thigmotaxis):
Respons op contact is gevarieerd bij Paramecium. Als het voorste uiteinde licht wordt aangeraakt met een fijne punt, treedt een sterke ontwijkende reactie op. Als een zwemmende Paramecium in botsing komt met een of ander voorwerp in het water, maar als het elders wordt aangeraakt, kan er geen reactie zijn. Een langzaam bewegend individu reageert vaak positief op contact met een voorwerp door er op te gaan liggen.
(ii) Reacties op chemicaliën (Chemotaxis):
Over het algemeen reageren Paramecia op een chemische prikkel door middel van het vermijden van reactie. Als een druppel zwakke zoutoplossing (0,5 procent) wordt ingebracht in een Paramecium-populatie op een microglijbaan, reageren de dieren met de vermijdende reactie en geen komt in de druppel. Op zuren daarentegen is de reactie positief, zelfs wanneer de concentratie sterk genoeg is om ze te doden.
(iii) Reacties op temperatuur (Thermotaxis):
Paramecium zoekt een optimale temperatuur van 24 tot 28°C. Bij een temperatuurverandering die duidelijk boven of onder het optimale bereik ligt, vertonen Paramecia een ontwijkende reactie. Een grotere warmte stimuleert snelle bewegingen en ontwijkende reacties totdat de dieren ontsnappen of gedood worden.
(iv) Reacties op licht (Phototaxis):
Met uitzondering van de groene Paramecium bursaria, die positief fototactisch is, zijn andere soorten onverschillig voor gewoon licht. Wanneer de lichtintensiteit echter plotseling en sterk wordt verhoogd, volgt over het algemeen een negatieve reactie. Paramecia vertonen een onmiddellijke negatieve reactie op ultraviolette stralen.
(v) Reacties op elektrische stroom (Galvanotaxis):
Paramecia reageren op elektrische prikkels. Wanneer twee elektroden tegenover elkaar worden geplaatst in een ondiepe schaal met Paramecia en een constante stroom wordt aangelegd, zwemmen alle organismen in dezelfde richting naar de kathode of negatieve elektrode, waar zij zich in grote aantallen concentreren.
Als de richting van de elektrische stroom wordt omgekeerd terwijl de Paramecia naar de kathode zwemmen, keren de organismen de richting om en zwemmen naar de nieuwe kathode.

(vi) Reacties op waterstroom (Rheotaxis):
Paramecia vertonen een positieve rheotaxis. In een zachte waterstroom zullen de Paramecia zich meestal met de stroom mee bewegen met hun voorste uiteinden stroomopwaarts.
(vii) Reacties op de zwaartekracht (Geotaxis):
Paramecia vertonen in het algemeen een negatieve reactie op de zwaartekracht, zoals te zien is in een kweek waarin veel individuen zich dicht onder de oppervlaktelaag verzamelen met hun voorste uiteinden naar boven. Als Paramecia in een omgekeerde met water gevulde U-vormige buis worden gebracht die aan beide uiteinden is afgesloten, bewegen zij zich onmiddellijk omhoog naar het horizontale deel van de buis.
Hervoortplanting bij Paramecium Caudatam:
Paramecium Caudatum plant zich aseksueel voort door transversale binaire splijting en ondergaat ook verschillende soorten nucleaire reorganisatie, zoals conjugatie, endomixis, autogamie, cytogamie en hemixis, enz.
(i) Transverse Binaire Splijting:
Transverse binaire splijting is de meest voorkomende vorm van aseksuele voortplanting bij Paramecium. Het is een duidelijk uniek aseksueel proces waarbij één volgroeid exemplaar zich in twee dochter-individuen deelt zonder een ouderlijk lijk achter te laten.
Het vlak van de deling loopt door het centrum van de cel en in een vlak dat loodrecht staat op de lange as van het lichaam. De deling van het cellichaam als geheel wordt altijd voorafgegaan door de deling van de kernen; het lijkt er zelfs op dat de voortplanting in gang wordt gezet door kernactiviteit en deling.

Paramecium Caudatum plant zich voort door transversale binaire splijting onder gunstige omstandigheden. Bij de binaire splijting deelt de micronucleus zich door mitose in twee dochter-micronuclei, die zich naar tegenovergestelde uiteinden van de cel verplaatsen. De macronucleus verlengt zich en deelt zich dwars door amitose.
Een andere cytofarynx wordt afgesneden en twee nieuwe contractiele vacuolen verschijnen, een bij het voorste uiteinde en een ander bij het achterste uiteinde. Intussen verschijnt er een insnoeringsgroef in het midden van het lichaam, die zich verdiept tot het cytoplasma volledig verdeeld is.
De resulterende twee “dochter”-paramecia zijn van gelijke grootte en bevatten elk een stel celorganellen. Van de twee geproduceerde dochterparamecia wordt de voorste proter genoemd en de achterste opisthe. Zij groeien uit tot volledige grootte voordat een nieuwe deling plaatsvindt.
Het proces van de binaire splitsing neemt ongeveer twee uur in beslag en kan één tot vier keer per dag plaatsvinden, waarbij 2 tot 16 individuen worden voortgebracht. In een jaar worden ongeveer 600 generaties geproduceerd.
De snelheid van de vermenigvuldiging hangt af van de externe omstandigheden van voedsel, temperatuur, leeftijd van de cultuur, en de bevolkingsdichtheid; ook van de interne factoren van erfelijkheid en fysiologie. Als alle nakomelingen van één individu zouden overleven en zich voortplanten, zou het aantal geproduceerde paramecia uiteraard al gauw gelijk zijn aan het volume van de aarde.
De term kloon wordt gebruikt om alle individuen aan te duiden die uit één individu zijn voortgekomen door splijting. Alle leden van een kloon zijn erfelijk gelijk.
(ii) Conjugatie:
Ordinair vermenigvuldigt Paramecium Caudatum zich door binaire splijting gedurende lange perioden, maar met tussenpozen kan dit worden onderbroken door het samenvoegen van twee dieren langs hun mondoppervlakken voor het sexuele proces van conjugatie.
Conjugatie wordt gedefinieerd als de tijdelijke vereniging van twee individuen die wederzijds microkernmateriaal uitwisselen. Het is een uniek type van een sexueel proces waarbij twee organismen zich scheiden kort na de uitwisseling van nucleair materiaal.
Sonneborn (1947) heeft op grond van paringsgedrag van Paramecium Caudatum gemeld dat elke soort Paramecium bestaat in een aantal variëteiten of syngens. Verder zijn er binnen elke syngens een aantal paringstypen, meestal twee.
De paringstypen blijven morfologisch identiek, maar zij vertonen fysiologische verschillen. Bij P. aurelia zijn er 14 syngens en 28 paringstypes, terwijl er bij P. caudatum 16 syngens en 32 paringstypes zijn. Er zijn waarnemingen gedaan dat paramecia zich gewoonlijk noch met leden van hun eigen paringstype noch met de andere variëteiten verenigen, maar alleen met het tweede paringstype van hun eigen variëteit.

Factoren die conjugatie induceren:
De factoren die conjugatie induceren verschillen van soort tot soort, maar enkele ervan worden hieronder gegeven:
1. Conjugatie treedt gewoonlijk op onder ongunstige levensomstandigheden; verhongering of voedseltekort en een bepaald bacterieel dieet of bepaalde chemicaliën zouden het proces van conjugatie bij bepaalde soorten Paramecium induceren.
2. Conjugatie treedt op na ongeveer 300 aseksuele generaties van binaire splijting, of het wordt afgewisseld met binaire splijting met lange tussenpozen om de stervende kloon te verjongen, d.w.z. het treedt op bij de individuen die het gewenste aantal aseksuele generaties moeten hebben doorgemaakt, de zogenaamde periode van onrijpheid, en dan worden zij geslachtsrijp om te conjugeren.
3. Conjugatie treedt op wanneer er een verandering optreedt in de fysiologische toestand van de paramecia, dan treedt zij op tussen zulke individuen die iets kleiner van omvang zijn (210 micron lang) en zij bevinden zich in een stadium dat kan worden beschouwd als een periode van ongezonde ouderdom; de paramecia van deze toestand zullen sterven indien zij niet de gelegenheid krijgen zich te conjugeren.
4. Plotselinge duisternis in lichtomstandigheden en lage temperaturen zouden bij sommige soorten het proces van conjugatie op gang brengen.
5. Conjugatie vindt niet plaats gedurende de nacht of duisternis; het begint in de vroege ochtend en gaat door tot in de middag.
6. Een eiwitachtige stof in de trilharen van paringstype individuen zou de conjugatie opwekken.
Proces van Conjugatie:
Het proces van conjugatie verschilt bij verschillende soorten Paramecium, maar de hierna volgende uiteenzetting heeft betrekking op het conjugatieproces van P. caudatum (Fig. 20.21).
In conjugatie komen twee Paramecium caudatum (pre-conjuganten genoemd) van de tegenovergestelde paringstypen van dezelfde soort met hun ventrale oppervlakken bij elkaar en verenigen zich door hun mondgroeven; hun cilia produceren een substantie op het lichaamsoppervlak die hechting van de twee conjugerende paramecia veroorzaakt.
Ze stoppen met eten en hun orale groef apparaat verdwijnt. De pellicula en het ectoplasma, op het punt van contact, van beiden breken af, en een protoplasmatische brug wordt gevormd tussen de twee dieren. Nu, deze individuen worden conjuganten genoemd.
In deze toestand zwemt het conjugerende paar actief en gelijktijdig vindt in elke conjugant een reeks nucleaire veranderingen plaats, zoals hieronder beschreven:
De macronucleus begint uiteen te vallen, hij wordt los van textuur en vormt een complexe gedraaide streng, tijdens de laatste helft van de conjugatieperiode zal hij tenslotte verdwijnen doordat hij in het cytoplasma wordt opgenomen. De micronucleus van elke conjugant deelt zich tweemaal, waarvan één reductiedeling.
Zo worden in elke conjugant vier haploïde dochter-micronuclei geproduceerd. Drie van deze vier micronuclei degenereren in elk, zodat er slechts één overblijft.
De resterende micronucleus van elke conjugant deelt zich mitotisch in twee ongelijke pronuclei of gametische kernen en vormt een grotere stationaire vrouwelijke pro-nucleus en een kleinere, actieve migrerende mannelijke pro-nucleus.
De migrerende pro-nucleus van de ene conjugant steekt de protoplasmabrug over en versmelt met de stationaire pro-nucleus van de andere conjugant om een synkaryon of conjugatienucleus te vormen waarin het diploïde aantal chromosomen is hersteld en er een uitwisseling van erfelijk materiaal heeft plaatsgehad.
Het proces is vergeleken met bevruchting bij hogere dieren, maar dit is geen bevruchting omdat er geen gameten bij betrokken zijn. De conjuganten scheiden zich nu (na ongeveer 12-48 uur) en worden ex-conjuganten genoemd. Het synkaryon van elke ex-conjugant deelt zich driemaal, zodat in elke ex-conjugant acht micronuclei worden gevormd.
Vier van de acht micronuclei vergroten zich en worden macronuclei, en drie van de andere vier micronuclei verdwijnen.
De overblijvende micronucleus deelt zich en tegelijkertijd deelt de ex-conjugant zich door binaire splijting in twee cellen, elk met twee macronuclei en één micronucleus. De cellen en hun micronuclei delen zich een tweede maal om uit elke ex-conjugant vier paramecia te vormen, zodat elk een macronucleus en een micronucleus heeft.
De nieuwe macronucleus, evenals de micronucleus, zijn gemaakt van nieuw materiaal. Deze nieuwe kernen bevatten waarschijnlijk nieuw en ander potentieel, dat bij de gezonde individuen tot uiting komt.
Betekenis van Conjugatie:
Een kloon zal uitsterven als nucleaire reorganisatie niet optreedt, maar de kloon kan worden verjongd om zijn vroegere kracht te herwinnen door nucleaire herschikking, deze nucleaire reorganisatie komt tot stand door conjugatie, dus, conjugatie is essentieel voor voortgezette binaire splitsing.
De betekenis van conjugatie is hieronder samengevat:
1. Conjugatie dient als een proces van verjonging en reorganisatie waardoor de vitaliteit van het ras wordt hersteld. Als er gedurende lange perioden geen conjugatie optreedt, verzwakken de paramecia en sterven zij af. (Woodruffs bewering dat hij paramecia gedurende 22.000 generaties zonder conjugatie gezond heeft gehouden, is door Sonneborn weerlegd, omdat hij aantoonde dat alle paramecia van Woodruff tot hetzelfde paringstype behoorden).
2. Er is geen geslachtsonderscheid in conjuganten, hoewel alleen paramecia van twee verschillende paringstypes van dezelfde variëteit zich zullen conjugeren.
3. Er is geen onderscheid naar geslacht, doch de actieve migrerende pro-nucleus wordt als mannelijk beschouwd en de stilstaande pro-nucleus als vrouwelijk.
4. Conjugatie is slechts een tijdelijke vereniging, er is geen versmelting van cytoplasma en er wordt geen zygote geproduceerd, maar de kern van elke ex-conjugant bevat erfelijk materiaal van twee conjugerende individuen.
5. Conjugatie brengt vervanging van de macronucleus door materiaal van het synkaryon teweeg, dit is een gebeurtenis van fundamenteel belang. Bij de binaire splitsing werden de chromosomen van de macronucleus willekeurig over de dochtercellen verdeeld, voortzetting van de binaire splitsing had de kloon zwak gemaakt met enkele structurele afwijkingen.
Conjugatie bewerkstelligt de vorming van het juiste aantal chromosomen in de macronucleus, zodat de race in kracht wordt vernieuwd. De rol van de micronucleus is het herstellen van een evenwichtig chromosomen- en genencomplex.
Aberrant gedrag bij de voortplanting in Paramecium Caudatum:
Paramecium Caudatum vertoont bepaalde afwijkingen in zijn nucleaire gedrag tijdens splitsing en conjugatie, deze afwijkingen zijn endomixis, autogamie, cytogamie en hemixis. Bij de eerste drie processen vindt genetische recombinatie plaats en wordt uit de micronucleus een nieuwe macronucleus gevormd.
(i) Endomixis:
Woodruff en Erdmann (1914) meldden als eersten een nieuw nucleair reorganisatieproces, endomixis (Gr., endon = binnen; mixis = vermenging) in Paramecium aurelia, een bimicronucleate soort (Fig. 20.22). Dit proces werd beschreven als een periodiek optredend proces waarbij een nieuw macronucleair apparaat wordt geproduceerd zonder synkaryonvorming. Endomixis treedt op binnen een enkel individu.
Volgens Woodruff en Erdmann degenereert de macronucleus en delen de micronuclei zich tweemaal om acht micronuclei te vormen. Zes micronuclei ontaarden en twee blijven over. Met nog slechts twee micronuclei over, deelt het dier zich door splijting in twee cellen, elk met één micronucleus.
De micronucleus van elke dochtercel deelt zich tweemaal om vier micronuclei te vormen. Twee micronuclei vergroten zich tot macronuclei. Het dier en zijn micronuclei delen zich, zodat twee dochter-individuen worden gevormd, elk met één macronucleus en twee micronuclei.
Endomixis komt voor in die variëteit van P. aurelia die niet conjugeert, vandaar dat het effect van endomixis hetzelfde kan zijn als dat van de conjugatie, aangezien beide processen vervanging van de macronucleus bewerkstelligen met materiaal van de micronucleus, en beide processen verjongen de vitaliteit van het ras.
Maar de twee processen verschillen omdat er bij endomixis geen versmelting van pronuclei plaatsvindt; endomixis kan worden vergeleken met parthenogenese.
Hoewel sommige werkers met goede redenen hebben beweerd dat endomixis geen geldig proces is en dat het is beschreven als gevolg van foutieve observatie. Naar alle waarschijnlijkheid vindt endomixis niet plaats en is het wellicht slechts een gespecialiseerd geval van autogamie.

Later hebben Erdmamm en Woodruff (1916) endomixis gemeld bij Paramecium caudatum. Diller gelooft echter niet in de geldigheid van dit proces en meent dat Erdmann en Woodruff eenvoudig de stadia van hemixis en autogamie hebben gecombineerd in één schema, endomixis.
(ii) Autogamie:
Diller (1934, 1936) en Sonneborn (1950) beschreven een proces van zelfbevruchting of autogamie dat zich voordoet in één enkel individu in Paramecium aurelia (Fig. 20.23). Hij meldde dat bij autogamie drie micronucleaire (pregamische) delingen, waarbij rijping optreedt, de gametische kernen (pronuclei) produceren.
Tijdens autogamie bij P. aurelia delen de twee micronuclei zich tweemaal (eenmaal meiotisch) om acht micronuclei te vormen, waarvan er zes degenereren. Ondertussen groeit de macronucleus uit tot een strengenachtige massa die later in stukken breekt om in het cytoplasma te worden opgenomen. Twee van de acht micronuclei gaan, als pronuclei, een protoplasmatische kegel binnen die bij de celmond uitpuilt.
De twee pronuclei versmelten tot synkaryon. Het synkaryon deelt zich twee keer en vormt vier micronuclei. Twee micronuclei worden macronuclei. Het Paramecium en zijn micronuclei delen zich en vormen twee dochter-individuen, elk met één macronucleus en twee micronuclei. Dit proces is in ongeveer twee dagen voltooid.
Autogamie brengt verjonging van het ras tot stand.
Het lijkt op conjugatie in zoverre dat de nieuwe macronucleus wordt gevormd door materiaal van de micronucleus, in de nieuwe macronucleus wordt het juiste aantal chromosomen hersteld; en ook in het feit dat fusie van twee pronuclei optreedt. Maar autogamie verschilt van conjugatie omdat slechts één individu deelneemt aan autogamie en het levert de beide pronuclei, het is een soort zelfbevruchting.
(iii) Hemixis:
Diller (1936) rapporteerde de hemixis bij Paramecium aurelia (Fig. 20.24). Hemixis is in de eerste plaats een proces van macro-nucleaire fragmentatie en deling zonder enige ongewone micro-nucleaire activiteit. Diller deelde hemixis in vier typen in, namelijk A, B, C, en D zoals in Fig. 20.24 in P. aurelia, maar hij trof ook alle typen aan in massaculturen van P. caudatum en P. multimicronucleatum.
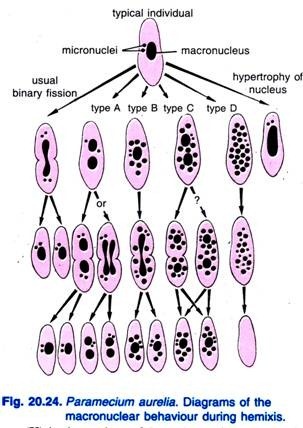
Type A is de eenvoudigste vorm van hemixis, die wordt gekenmerkt door een splitsing van de macronucleus in twee of meer delen. Deze deling loopt niet synchroon met de deling van de micronucleus.
Type B wordt gekenmerkt door de extrusie van één tot 20 of meer chromatinebolletjes uit de macronucleus in het cytoplasma.
Type C wordt gekenmerkt door de gelijktijdige splitsing van de macronucleus in twee of meer grote delen en de extrusie van macronucleusbolletjes in het cytoplasma.
Type D wordt geacht pathologische condities te vertegenwoordigen waarbij de macronucleus volledige fragmentatie ondergaat in chromatinebolletjes die uiteindelijk uit de cel verdwijnen. Micronuclei verdwijnen in het algemeen vóór de ontbinding van de macronucleus.
(iv) Cytogamie:
Wichterman (1939) heeft een ander sexueel proces bij Paramecium caudatum gerapporteerd, dat hij cytogamie heeft genoemd. Bij cytogamie is er geen uitwisseling van kernen. Bij dit proces komen twee individuen aan hun ventrale oppervlakken bij elkaar, maar de pellicula van de twee individuen breekt niet af.
De micronucleus van elk individu deelt zich driemaal om acht micronuclei te vormen, waarvan er zes in elk individu uiteenvallen. De twee overblijvende micronuclei versmelten tot een synkaryon in elke cel. De dieren scheiden zich nu.
Cytogamie verschilt van autogamie in die zin dat er twee dieren met elkaar in contact zijn, maar het lijkt op autogamie en conjugatie in de fusie van twee pronuclei. Cytogamie verschilt van conjugatie in die zin dat er geen nucleaire uitwisseling plaatsvindt tussen de twee dieren die bij elkaar komen.
Enkele Cytoplasmatische Deeltjes Gemeld in Paramecium Caudatum:
(i) Kappa Deeltjes:
In 1938 rapporteerde T.M. Sonneborn dat sommige rassen (bekend als killers of moordenaarsstammen) van Paramecium een giftige stof produceren, paramecine genaamd, die dodelijk is voor andere individuen, sensitieven genaamd. Het paramecine is oplosbaar in water, verspreidbaar en voor zijn produktie afhankelijk van enkele deeltjes die zich in het cytoplasma van de Paramecium (moordenaarsstam) bevinden.
Deze deeltjes worden kappa-deeltjes genoemd. De kappa deeltjes bevatten DNA en RNA. Een killer Paramecium kan honderden kappa deeltjes bevatten. Uit de gedetailleerde bestudering van deze deeltjes is gebleken dat een dominant gen (K) in de kern van Paramecium noodzakelijk is voor het bestaan van kappa-deeltjes, voor hun vermenigvuldiging en voor de productie van paramecine.
(ii) mµ, Deeltjes:
R.W. Siegel (1952) rapporteerde een ander type mp-deeltjes in het cytoplasma van sommige Paramecium. Een Paramecium met mp partikels wordt mate killer genoemd, omdat wanneer het zich verbindt met een Paramecium zonder mµ partikels, mate sensitive genaamd, dan doodt het de laatste. De mp-deeltjes zijn ook samengesteld uit DNA, RNA, enz.
Deze deeltjes bestaan alleen in die parameciums waarvan de micronucleus ten minste één dominant gen bevat van een van de twee paren niet-gekoppelde chromosomale genen (M1 en M2).
(iii) Pi-deeltjes:
Deze deeltjes worden verondersteld de gemuteerde vorm van kappa-deeltjes te zijn, maar zij produceren geen enkel soort giftige stof.
(iv) Lambda Deeltjes:
Deze deeltjes worden gemeld in killer paramecia en zouden een of andere stof produceren die verantwoordelijk is voor het veroorzaken van lysis of desintegratie van gevoelige paramecia, d.w.z., die deze niet bezitten.