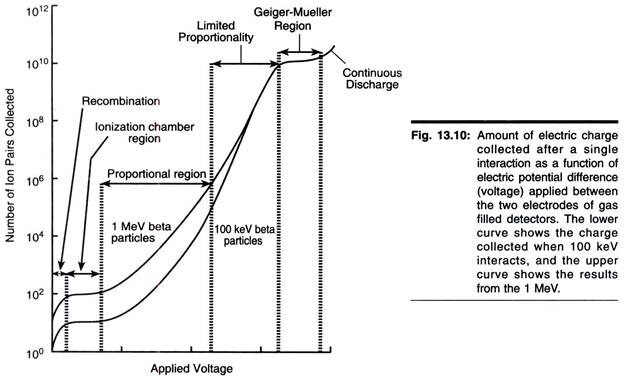
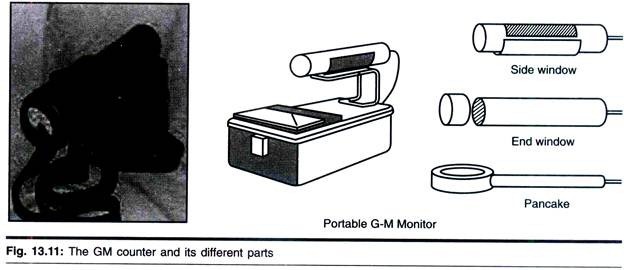
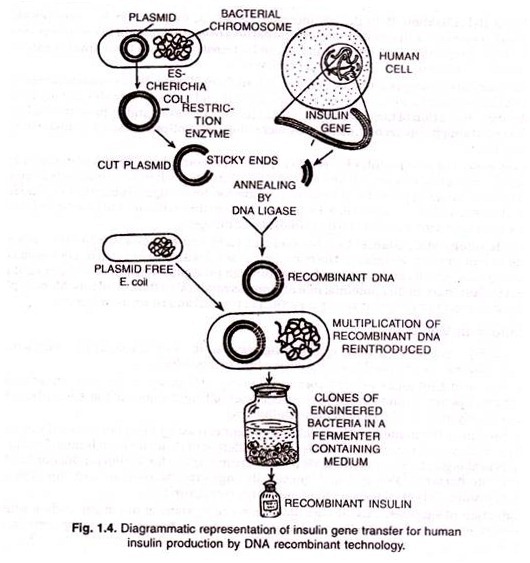
ADVERTISEMENTS:
W tym artykule będziemy dyskutować o Paramecium Caudatum:- 1. Habitat, siedlisko i kultura Paramecium Caudatum 2. Struktura Paramecium Caudatum 3. Lokomocja 4. Odżywianie 5. Oddech i wydalanie 6. Osmoregulacja 7. Zachowanie 8. Reprodukcja 9. Aberracyjne zachowania w rozmnażaniu 10. Niektóre cząstki cytoplazmatyczne.
Zawartość:
- Habit, Habitat i kultura Paramecium Caudatum
- Struktura Paramecium Caudatum
- Lokomoc Paramecium Caudatum
- Odżywianie w Paramecium Caudatum
- Oddychanie i wydalanie w Paramecium Caudatum
- Osmoregulacja Paramecium Caudatum
- Behaviour of Paramecium Caudatum
- Reproduction in Paramecium Caudatum
- Aberrant Behaviour in Reproduction in Paramecium Caudatum
- Some Cytoplasmic Particles Reported in Paramecium Caudatum
- Habit, Habitat i hodowla Paramecium Caudatum:
- Kultura Paramecium:
- Struktura Paramecium Caudatum:
- (i) Wielkość i kształt Paramecium Caudatum:
- (ii) Otoczka:
- (iii) Cilia:
- (iv) Układ rzęskowy:
- (v) Oral Groove i Cytopyge:
- (vi) Cytoplazma:
- (vii) Ektoplazma:
- (viii) Trichocysty:
- (ix) Neuromotorium i związane z nim fibryle:
- (x) Endoplazma:
- (xi) Wakuole kurczliwe:
- (xii) Wakuole pokarmowe:
- Lokomoc Paramecium Caudatum:
- (i) Metaboly lub Body Contortions:
- (ii) Ruch rzęskowy:
- Odżywianie Paramecium Caudatum:
- Mechanizm odżywiania:
- Trawienie i połykanie Paramecium Caudatum:
- Wdychanie i wydalanie Paramecium Caudatum:
- Zachowanie Paramecium Caudatum:
- (i) Reakcje na kontakt (Thigmotaxis):
- (ii) Reakcje na substancje chemiczne (chemotaksja):
- (iii) Reakcje na temperaturę (termotaksja):
- (iv) Reakcje na światło (fototaksja):
- (v) Reakcje na prąd elektryczny (galwanotaksja):
- (vi) Reakcje na prąd wody (reotaksja):
- (vii) Reakcje na grawitację (Geotaksja):
- Reprodukcja u Paramecium Caudatam:
- (i) Poprzeczne rozszczepienie binarne:
- (ii) Koniugacja:
- Zachowanie jądrowe w reprodukcji u Paramecium Caudatum:
- (i) Endomiksja:
- (ii) Autogamia:
- (iii) Hemixis:
- (iv) Cytogamia:
- Niektóre cząsteczki cytoplazmatyczne zgłoszone w Paramecium Caudatum:
- (i) Cząsteczki Kappa:
- (ii) mµ, cząsteczki:
- (iii) Cząsteczki Pi:
- (iv) Cząstki lambda:
Habit, Habitat i hodowla Paramecium Caudatum:
Paramecium caudatum (Gr., paramekes = podłużny; L., caudata = ogon) jest powszechnie spotykany w słodkowodnych stawach, basenach, rowach, strumieniach, jeziorach, zbiornikach i rzekach. Szczególnie obficie występuje w stagnujących stawach bogatych w rozkładającą się materię, w zalewach organicznych oraz w wodach ściekowych. Paramecium caudatum jest organizmem wolno żyjącym, a jego zasięg występowania jest ogólnoświatowy.
Kultura Paramecium:
ADVERTISEMENTS:
Zabierz zanurzone chwasty ze stawu i umieść w słoiku z wodą destylowaną, przykryj słoik i pozostaw do zgnicia; roje Paramecia pojawią się w ciągu kilku dni. Teraz ugotuj siano w wodzie, odcedź wywar i dodaj kilka ziaren pszenicy, i pozostaw do czasu, aż zmętnieje od bakterii.
Przenieś Paramecia z pierwszego słoika do tego płynu, gdzie będą się szybko mnożyć. Same napary z siana wytworzą Paramecia wykazujące obecność cyst, a cysty przypominające ziarna piasku zostały zgłoszone, ale nie ma dowodu na to, że Paramecia tworzą cysty, ponieważ nigdy nie zostały potwierdzone.
Struktura Paramecium Caudatum:
(i) Wielkość i kształt Paramecium Caudatum:
Paramecium caudatum (rys. 20.1) jest organizmem mikroskopijnym i widocznym gołym okiem jako drobne wydłużone ciało.
ADWERSJE:
Wygląda na jasnoszare lub białe, mierząc powszechnie od 170 do 290 mikronów długości i może osiągnąć długość do 300-350 mikronów. P. caudatum wygląda jak podeszwa pantofla lub buta, stąd zwierzę to jest powszechnie znane jako slipper animalcule. Jest czterokrotnie dłuższy od szerokiego i nieco cylindryczny z wyraźnie odmiennymi końcami.
Przód poruszająca się przednia część jest smukła z tępym lub zaokrąglonym końcem, podczas gdy tylny koniec jest nieco spiczasty lub stożkowaty. Najszersza część organizmu znajduje się tuż poniżej środka. Ciało zwierzęcia jest asymetryczne w formie, z dobrze zaznaczoną powierzchnią ustną lub brzuszną i brzuszną lub grzbietową.
(ii) Otoczka:
Ciało pokryte jest cienką, dwuwarstwową, elastyczną i twardą otoczką wykonaną z żelatyny. Otoczka utrzymuje kształt zwierzęcia, ale jest wystarczająco elastyczna, aby umożliwić skurcze. Otoczka ma podwójną błonę, zewnętrzna błona jest ciągła z rzęskami, a wewnętrzna z ektoplazmą. Pod większym powiększeniem mikroskopu na powierzchni otoczki widoczne są prostokątne lub sześciokątne wgłębienia.

Układ ten jest zachowany na powierzchni grzbietowej Paramecium, ale na powierzchni brzusznej grzbiety zbiegają się z przodu i z tyłu w kierunku otworu przedoczodołowego i otworu tylnego. Każde sześciokątne wgłębienie jest przedziurawione centralnym otworem, przez który wychodzi pojedyncze cilium. Na przednich i tylnych krawędziach sześciokątnych wgłębień znajdują się otwory trichocyst.
Mikroskopowe badanie elektronowe otoczki (ryc. 20.2) przez Ehret i Powersa (1957) ujawniło, że sześciokątne zagłębienia odpowiadają regularnym seriom jamek, pęcherzyków płucnych. Wszystkie pęcherzyki płucne tworzą razem ciągłą warstwę pęcherzykową, która jest ograniczona przez zewnętrzną i wewnętrzną błonę pęcherzykową.
Warstwa zewnętrzna leży w ścisłym kontakcie pod błoną komórek zewnętrznych. Dlatego w skład pecherzyka wchodzi zewnętrzna błona komórkowa, zewnętrzna błona pęcherzykowa i wewnętrzna błona pęcherzykowa.
(iii) Cilia:
Całe ciało pokryte jest licznymi, drobnymi, przypominającymi włoski występami zwanymi rzęskami. Cilia występują w podłużnych rzędach na całym ciele, warunek ten jest znany jako holotrichous w którym rzęski ciała są równe. Cilia mają taką samą strukturę jak flagella, mają zewnętrzną osłonę protoplazmatyczną lub błonę plazmatyczną z dziewięcioma podwójnymi podłużnymi włóknami w obwodowym pierścieniu. W niektórych rzęskach dziewięć zewnętrznych fibrylli nie jest sparowanych.
WSKAZANIA:
Dwie środkowe podłużne fibryle są cieńsze niż fibryle zewnętrzne. Każde cilium powstaje z bazalnej granulki lub kinetosomu. Dziewięć par fibryli obwodowych łączy się razem, tworząc ścianę kinetosomu, tak więc kinetosom jest rurką, która jest albo otwarta albo zamknięta w swoim dolnym końcu, dwie centralne fibryle zatrzymują się na poziomie otoczki u większości rzęsistków.
Wyrastający z kinetosomu jest cienki rizoplast, który nie łączy się z jądrem. Wiele Metazoa ma również rzęski, ich struktura jest taka sama, z wyjątkiem tego, że ziarnistość podstawna jest inna i ma drobne włókna lub włókna korzeniowe sięgające w dół do cytoplazmy. Ale rzęski różnią się od flagelli tym, że są na ogół liczniejsze i krótsze.
Ciliatura może być wygodnie podzielić na ciało lub somatyczne rzęsek, które znajdują się na powierzchni ciała, i do ustnej ciliature, która jest związana z regionu ust. Ciało rzęski są równe, ale są one dłuższe na tylnym końcu, stąd nazwa caudatum. Rzęski są organellami lokomocji i pobierania pokarmu, pełnią też rolę receptorów czuciowych i wykrywają bodźce środowiska zewnętrznego.

Ultra struktura rzęsek:
Cilium i flagella mają skład fibrylarny. U podstawy rzęski mają średnicę około 0,2 mikrona lub 2,000 A0, która może wynosić do 10 mikronów ponad powierzchnię komórki. Cylindry są ograniczone jednostkową błoną o grubości 90 A0, która przypomina błonę plazmatyczną i pozostaje w ciągłości z nią. W przestrzeni ograniczonej rzęskami znajduje się wodnista substancja zwana macierzą.
W macierzy pozostaje osadzonych jedenaście podłużnych fibryli lub mikrotubul. Z jedenastu fibryli, dwie znajdują się w centrum, podczas gdy pozostałe dziewięć fibryli pozostaje ułożone peryferyjnie wokół centralnych fibryli. Każda z dziewięciu zewnętrznych fibryli ma średnicę 360A0 i składa się z dwóch subfibryli o średnicy od 180 do 250A0.
Te podfibryle są oznaczone jako podfibryle A i podfibryle B.
ZALECENIA:
Podwłókno A jest nieco większe niż podwłókno B. Podwłókno A wydaje dwa grube występy lub ramiona z jednej strony. Ramiona subfibryli A wszystkich zewnętrznych fibrylli pozostają skierowane w kierunku zgodnym z ruchem wskazówek zegara. Ponadto, podwłókno A występuje bliżej środka cilium niż podwłókno B. Oba podwłókna mają wspólną ścianę o grubości 50A0.
Dwa centralne fibryle nie mają sparowanych podfibryli jak peryferyjne dziewięć fibryli, ale każda zawiera tylko pojedynczą rurkę. Każdy centralny fibryl ma średnicę około 250A0 i składa się z 60A0 grubej ściany.
Oba centralne fibryle pozostają oddzielone przez przestrzeń 350A0 i pozostają otoczone wspólną powłoką. Gibbnos (1967) podał, że otoczka centralnych fibrylli wydaje dziewięć promieniście zorientowanych ogniw lub szprych do każdego podfibryla A.

Mikroskopia elektronowa wysokiej rozdzielczości ujawniła, że każda z obwodowych i centralnych fibrylli rzęsek i flagelli składa się z dziesięciu do dwunastu włókien o grubości 40A0. Każde włókno jest paciorkowate. Każdy paciorek jest ułożony w siatce 40 na 50A0 w płaszczyźnie ściany kanalika. Te koraliki są uważane za podstawową podjednostkę struktury kanalika.
(iv) Układ rzęskowy:
Układ infra rzęskowy znajduje się tuż poniżej pęcherzyków płucnych. Składa się on z kinetosomu lub ciała podstawowego i kinetodesma. Rzęski powstają z kinetosomów, a z każdego kinetosomu wyrastają delikatne włókna cytoplazmatyczne zwane kinetodesmą, (ryc. 20.2).
Leżąca poniżej pecherzyka nieco na prawo, ale połączona ze wszystkimi kinetosomami jednego podłużnego rzędu, jest podłużną wiązką kilku kinetodesmatów, kinetodesmata każdego kinetosomu rozciąga się na odległość przednią do własnej wiązki kinetodesmatów.
Podłużny rząd kinetosomów z ich kinetodesmatami tworzy podłużną jednostkę zwaną kinetą. Wszystkie kinety lub kinetia tworzą system infra-ciliary u rzęsek. Kinetia leżą w korze pod otoczką, ich liczba jest dość stała u każdego rzęsistka.
System infra-ciliary kontroluje i koordynuje ruchy rzęsek, i doprowadza do powstania organelli w podziale komórkowym, np. niektóre kinetia tworzą usta. W rozszczepieniu binarnym u rzęsistków kinetia są przecięte poprzecznie na dwie części, z których każda przechodzi do jednej komórki potomnej, jest to tzw. rozszczepienie perikinetalne.
(v) Oral Groove i Cytopyge:
Na stronie brzuszno-bocznej znajduje się duże skośne, płytkie wgłębienie zwane rowkiem ustnym lub perystomem, które nadaje zwierzęciu asymetryczny wygląd. Biegnie on skośnie do tyłu z jednej strony (zwykle z lewej na prawą, ale w niektórych przypadkach z prawej na lewą) i kończy się nieco za środkową częścią ciała. Bruzda ustna prowadzi do krótkiego stożkowatego zagłębienia w kształcie lejka, zwanego przedsionkiem.
WSKAZANIA:
Przedsionek prowadzi bezpośrednio do nieruchomego, owalnego otworu zwanego cytostomem (jamą gębową). Bezpośrednio od cytostomu w kierunku środka ciała rozciąga się szeroka cytofarynka. Następnie cytofarynka skręca ostro w kierunku tylnej strony, stając się smukłym, zwężającym się przełykiem.
Tak więc, przełyk jest w przybliżeniu równoległy do powierzchni ciała Paramecium, z wyjątkiem jego tylnego krańca. Tutaj przełyk skręca ponownie w kierunku centrum zwierzęcia, aby doprowadzić do tworzącej się wakuoli pokarmowej.
Cytopyge (również określane jako anus komórek lub punkt odbytu lub cytoproct) leży na brzusznej powierzchni ciała prawie pionowo za cytostomu lub usta. Niestrawione cząstki pokarmu są eliminowane przez cytopeptyd. Ciliation cytopharynx jest bardzo skomplikowana.
Gelei (1934) poinformował o obecności czterech rzędów i Lund (1941) obserwowane co najmniej cztery rzędy. Struktura zwana penniculus znajduje się na lewej ścianie cytopharynx i spiralnie przechodzi przez około 90 stopni tak, że jej tylny koniec znajduje się na ustnej (brzusznej) powierzchni przełyku.
Według Lunda, penniculus składa się z ośmiu rzędów rzęsek ułożonych w dwóch ściśle ustawionych bloków każdego. Podobne pasmo składające się z czterech rzędów długich rzęsek, które są mniej zwarte niż w penniculus jest określany jako quadrulus. Spiralnie biegnie ono w dół grzbietowej ściany jamy policzkowej i kończy się blisko penniculus.
Penniculus i quadrulus zostały błędnie nazwane falistą membraną przez niektórych pracowników. Quadrulus i penniculus kontrolować przejście żywności. Nie wiadomo jak działają rzęski, prawdopodobnie ich włókna kurczą się w sposób rytmiczny, co powoduje zginanie. Gelei (1925) zwrócił uwagę, że funkcją penniculusa jest wtłaczanie elementów pokarmowych do organizmu.

(vi) Cytoplazma:
Cytoplazma jest zróżnicowana na wąską, zewnętrzną lub korową strefę zwaną ektoplazmą i większy, wewnętrzny lub śródplazmatyczny region zwany endoplazmą.
(vii) Ektoplazma:
Ektoplazma (ektosarc lub cortex) jest stałą częścią ciała, wyraźnie odgraniczoną od endoplazmy. Ektoplazma tworzy twardą, przejrzystą, cienką i gęstą warstwę zewnętrzną. Zawiera ona trichocysty, rzęski i struktury fibrylarne, a na zewnątrz jest ograniczona osłoną zwaną pelliką.
(viii) Trichocysty:
Umieszczone w ektoplazmie pod kątem prostym do powierzchni są małe wrzecionowate torebki zwane trichocystami. Mała plamka na każdym przednim i tylnym brzegu sześciokąta oznacza położenie trichocysty. Są one wypełnione refrakcyjnym, gęstym płynem mającym substancję pęczniejącą, na zewnętrznym końcu znajduje się stożkowata główka lub kolec.
Trychocysty leżą prostopadle w ektoplazmie, otwierają się małymi porami na grzbietach sześciokątnych obszarów otoczki.
Wyrastają z kinetosomów rzęsek, następnie migrują i lokują się w równej odległości w endoplazmie. Gdy zwierzę jest podrażnione, trichocysty są uwalniane w postaci długich, lepkich nici. Uwolniona trichocysta ma nieprzezroczysty kolec przypominający odwrócony paznokieć i długi prążkowany trzon, ale trzon nie jest widoczny w stanie nieuwolnionym i prawdopodobnie powstaje podczas uwalniania.
Funkcja trichocyst jest niepewna, ale są one uwalniane jako reakcja na lokalne kontakty i urazy, mogą służyć jako organelle obronne.
Ale to jest niepewne, ponieważ trichocysty są nieskuteczne wobec Didinium, głównego drapieżnika na Paramecium, mogą służyć do mocowania zwierzęcia do miejsca podczas karmienia. U niektórych rzęsistków trichocysty działają jako organella ofensywne. Po wypuszczeniu trichocyst, regenerują się z kinetosomów.


(ix) Neuromotorium i związane z nim fibryle:
Według Lunda (1933) na lewej grzbietowej ścianie cytofarynki na poziomie tylnego brzegu cytostomu znajduje się bardzo mała, dwuboczna masa, neuromotorium. Z neuromotorium włókna promieniują do endoplazmy.
Z tych czterech lub więcej zazwyczaj przechodzą prawie do grzbietowej ściany ciała, ale reszta jest krótsza i nie jest określona w pozycji. Wszystkie są określane jako fibryli endoplazmatycznych. Ich funkcja jest nieznana, ale mogą one koordynować ruchy żywieniowe rzęsek ustnych. Włókna mogą również zapewniać wsparcie mechaniczne, elastyczność, kurczliwość, przewodnictwo i wpływ na metabolizm.

(x) Endoplazma:
Endoplazma lub medulla jest bardziej płynną i obszerną częścią cytoplazmy, która zawiera wiele ziarnistości cytoplazmatycznych, jak również inne inkluzje i struktury o wyspecjalizowanym charakterze. Inkluzje cytoplazmatyczne to mitochondria, aparaty Golgiego, wakuole, kryształy, granulki i chromidia, itp. W endoplazmie znajdują się również inne struktury, takie jak jądra, wakuole kurczliwe i wakuole pokarmowe.
Jądra:
W endoplazmie w pobliżu cytostomu znajdują się dwa jądra, tj. Paramecium jest heterokariotyczne, duże elipsoidalne i ziarniste makrojądro oraz inne małe zwarte mikrojądro.
Makronukleus jest widocznym, elipsoidalnym lub nerkowatym ciałem. Jest to typ zwarty, zawierający cienkie nici i ciasno upakowane dyskretne ziarna chromatyny o zmiennej wielkości, osadzone w achromatycznej matrycy. Posiada wiele nukleoli i znacznie więcej materiału chromatynowego (DNA).
Jest to jądro somatyczne lub wegetatywne. Dzieli się amitotycznie i kontroluje funkcje wegetatywne (czynności metaboliczne) zwierzęcia. Poddaje się mitozie.
Jądro mikronuklearne jest małe, zwarte i kuliste. Zwykle znajduje się w pobliżu makrojądra, często we wklęsłości. Drobne ziarna i nici chromatyny są równomiernie rozmieszczone w całej strukturze. Mikronukleus dzieli się mitotycznie i kontroluje rozmnażanie.
Liczba mikrojąderek zmienia się w zależności od gatunku; jest to jedno u P. caudatum, dwa u P. aurelia i wiele u P. multimicronucleatum. Mikrojądro zawiera wyraźny nukleus u P. aurelia, ale nie występuje on u P. caudatum. Moses (1949; 1950) stwierdził, że makrojądro i mikrojądro są identyczne pod względem składu chemicznego.
(xi) Wakuole kurczliwe:
Występują dwie duże, wypełnione płynem wakuole kurczliwe, każda położona w pobliżu jednego końca ciała blisko powierzchni grzbietowej. Ich położenie jest stałe (w przeciwieństwie do ameby), leżą między ektoplazmą a endoplazmą, ale są to organelle tymczasowe, okresowo zanikające. U niektórych gatunków wydają się mieć błonę wyściełającą, w którym to przypadku nie znikają całkowicie podczas skurczu.
Połączone z każdą wakuolą kurczliwą są pięć do dwunastu rurkowatych promieniujących kanałów, z których każdy składa się z części końcowej, długiej ampulli, która zapada się, gdy jest pusta, i krótkiego kanału iniekcyjnego, który otwiera się do wakuoli.

Kanały komunikują się z dużą częścią ciała, skąd pobierają płyny i wlewają je do wakuoli, która w ten sposób odtwarza się i zwiększa swoje rozmiary, kiedy wakuola kurczliwa osiąga swój maksymalny rozmiar, kurczy się nagle (skurcz) i wypuszcza swoją zawartość przez stały otwór w błonce, wtedy kanały ponownie tworzą wakuole kurczliwe, kanały nie znikają całkowicie, ponieważ są strukturami trwałymi.
Dwie wakuole kurczliwe rozładowują się nieregularnie, tylna kurczy się szybciej, ponieważ znajduje się w pobliżu cytofarynki i dostaje się do niej więcej wody. Główną funkcją kanałów i wakuoli kurczliwych jest funkcja hydrostatyczna, usuwają one nadmiar wody z protoplazmy, woda jest częściowo wchłaniana, a częściowo pobierana podczas odżywiania.
Zazotowa substancja odpadowa zawiera związki amoniaku i trochę moczanów, które są wydalane z wakuoli kurczliwych wraz z CO2, ale nie ma dowodów na to, że substancja wydalnicza jest wydzielana przez protoplazmę do kanałów lub wakuoli kurczliwych.
Poniżej perystomu, po jednej stronie cytofarynki, znajduje się stała cytoplazma lub odbyt, przez który wydalane są niestrawione resztki pokarmu i moczany.
Badanie mikroskopem elektronowym wakuoli kurczliwych wykazało, że każdy aparat kurczliwy składa się z niektórych kanalików retikulum endoplazmatycznego, kanalików nefrydialnych, kanałów doprowadzających, wakuoli pomocniczych (kanały promieniste) i głównej wakuoli kurczliwej. Przypuszcza się, że wakuole akcesoryjne są ampułkami kanalików doprowadzających (ryc. 20.11).
(xii) Wakuole pokarmowe:
Są to w przybliżeniu kuliste, niekurczliwe ciała różniące się wielkością i liczbą, leżące w endoplazmie. Zawierają one spożyte cząstki żywności, głównie bakterie i niewielką ilość płynu ograniczonego przez cienką błonę definitywną. Volkonsky (1934) zaproponował nazwę gastriole dla tych wakuoli. Związane z wakuolami żywności są granulki trawienne.
Lokomoc Paramecium Caudatum:
Paramecium Caudatum wykonuje lokomocję za pomocą dwóch metod, viz., metabolizm lub kontury ciała i przez rzęski.
(i) Metaboly lub Body Contortions:
Ciało Paramecium Caudatum posiada elastyczność, może przecisnąć się przez przejście węższe niż jego ciało, po czym ciało przyjmuje swój normalny kształt. Ta tymczasowa zmiana kształtu ciała jest metaboliczna, jest wywoływana w Paramecium przez protoplazmę.
(ii) Ruch rzęskowy:
Lokomoc wywołana przez rzęski jest główną metodą. Rzęski mogą bić do przodu lub do tyłu, umożliwiając zwierzęciu pływanie w kierunku przednim lub tylnym.
Normalnie zwierzę pływa do przodu, rzęski biją do tyłu, ale ukośnie, rzęski sztywnieją i wyginają się gwałtownie do tyłu, aby prawie dotknąć powierzchni ciała, to się nazywa skuteczny udar; następnie rzęski stają się wiotkie i wracają powoli do pierwotnej pozycji pionowej, to się nazywa udar odzysku.
Łąkotki tego samego rzędu poprzecznego biją razem, a rzęski tego samego rzędu podłużnego biją jedna za drugą od przedniego do tylnego końca.

Ten skoordynowany ruch rzęsek nazywamy rytmem metachronicznym, który zawdzięczamy układowi rzęskowemu wewnętrznemu; powoduje on pływanie zwierzęcia do przodu. Ale kiedy rzęski ciała biją ukośnie do tyłu, to w tym samym czasie dłuższe rzęski rowka ustnego biją bardziej energicznie, co powoduje, że przedni koniec odchyla się w lewo.
Działanie rzęsek ciała i bruzdy ustnej powoduje obrót zwierzęcia wokół jego długiej osi. Obrót ten jest zawsze w lewo (z wyjątkiem P. calkinsi, który obraca się w prawo).
Ta kombinacja ruchu do przodu, zawracania i rotacji sprawia, że zwierzę porusza się do przodu po spiralnej ścieżce w kierunku przeciwnym do ruchu wskazówek zegara. Tor ten ma prostą oś, a ta sama powierzchnia ciała zwierzęcia pozostaje w kierunku osi toru spiralnego. Natomiast podczas pływania do tyłu, wszystkie gatunki obracają się w prawo.
Bicie rzęsek może być odwrócone tak, że rzęski poruszają się skośnie do przodu, przez co zwierzę pływa do tyłu. Dzięki działaniu rzęsek, Paramecium porusza się z prędkością 1500 mikronów lub nawet więcej na sekundę.
Jennings twierdzi, że spirala Paramecium wynika z faktu, że podczas gdy rzęski uderzają głównie do tyłu, robią to ukośnie w prawo powodując tym samym przewrócenie się zwierzęcia na lewą stronę.
Also to kołysanie ciała w kierunku powierzchni aboralnej jest w dużej mierze ze względu na większą moc skutecznego skoku rzęsek ustnych, które uderzają bardziej bezpośrednio do tyłu. Rezultat – obrót Paramecium wokół długiej osi – umożliwia Paramecium podążanie mniej lub bardziej prostym kursem podczas tworzenia dużych spiral.
Odżywianie Paramecium Caudatum:
W Paramecium Caudatum, odżywianie jest holozoiczne. Pokarm składa się głównie z bakterii i drobnych Protozoa. Paramecium nie czeka na pokarm, ale aktywnie na niego poluje.
Twierdzi się, że Paramecium Caudatum wykazuje wybór w doborze pokarmu, ale wydaje się, że nie ma podstaw do tego, chociaż pochłania tylko niektóre rodzaje bakterii; dostępne dane sugerują, że 2 do 5 milionów jednostek Bacillus coli są pożerane przez jednego Paramecium w ciągu 24 godzin. Żywi się również jednokomórkowymi roślinami, takimi jak algi, okrzemki, itp. oraz małymi kawałkami zwierząt i warzyw.
Mechanizm odżywiania:
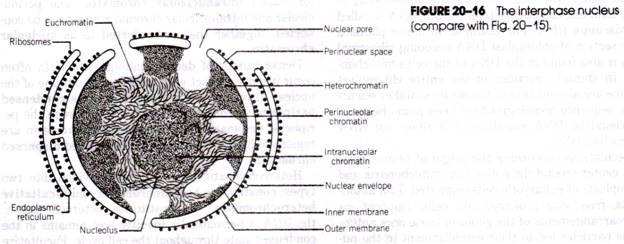
Kiedy Paramecium Caudatum wchodzi w rejon obfitego pożywienia, przychodzi na spoczynek. Żywi się tylko w spoczynku lub gdy pływa bardzo powoli, nigdy nie żywi się, gdy pływa szybko. Bicie rzęsek rowka ustnego powoduje, że stożkowaty wir wody pełnej pokarmu jest wciągany do rowka ustnego z pewnej odległości przed przednim końcem (rys. 20.16).
Cząstki pokarmu przechodzą następnie do przedsionka, skąd niektóre cząstki pokarmu są odrzucane i wyrzucane na zewnątrz, ale inne przechodzą do cytostomu.
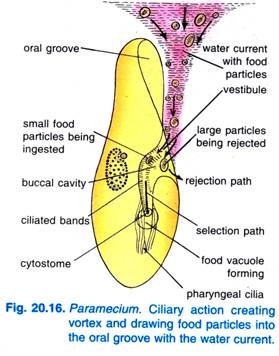
Na końcu cytofarynki powstaje wakuola pokarmowa, która wypełnia się cząstkami pokarmu. Quadrulus i peniculi kontrolują przechodzenie pokarmu do wakuoli pokarmowej, która tworzy się bocznie. Kiedy wakuola żywności osiąga pewną wielkość włókna post-buccal zacisnąć wakuolę żywności i to jest pinched off przez nich i zaczął na swój kurs.
Wakuola zawiera trochę wody oprócz żywności. Rotacyjne ruchy strumieniowe endoplazmy nazwie cyklozy nosić wakuoli żywności wzdłuż określonego kursu, który jest funkcjonalnie równoważne do przewodu pokarmowego.
Trakt zaczyna się od końca cytopharynx, a następnie do tylnej strony, a następnie do przodu, aby krążyć z endoplazmy, a następnie do powierzchni grzbietowej, a następnie w kierunku przedniego końca, a następnie w dół do cytopyge. Na początku swojej drogi wakuola pokarmowa zmniejsza swój rozmiar, potem znowu zwiększa.

Trawienie i połykanie Paramecium Caudatum:
Podczas cyklozy trawienie zachodzi dzięki enzymom wydzielanym przez protoplazmę do wakuoli. W procesie trawienia białka są przekształcane w aminokwasy, węglowodany w rozpuszczalne cukry i glikogen, prawdopodobnie trawione są również tłuszcze.
Zawartość wakuoli żywności są początkowo kwaśne (pH około 4), a następnie stają się zasadowe, główne trawienie występuje podczas fazy zasadowej. Niestrawiona materia jest z pewną siłą wyrzucana przez cytoplazmę.
Cykloza może być zademonstrowane doświadczalnie; jeśli mleko barwione czerwienią Kongo jest podawany do Paramecium, kuleczki tłuszczu mleka w wakuolach żywności najpierw zmienić kolor na czerwony z powodu kwaśnej reakcji enzymów, a następnie będą one zmienić z odcieni fioletu do niebieskiego z powodu reakcji alkalicznej, wakuole pokażą przebieg cyklozy.
Wdychanie i wydalanie Paramecium Caudatum:
Wymiana gazów (tlenu i dwutlenku węgla) odbywa się przez półprzepuszczalną otoczkę, podobnie jak u innych pierwotniaków słodkowodnych, w procesie dyfuzji. Paramecium Caudatum pozyskuje tlen z otaczającej go wody. Dwutlenek węgla i odpady organiczne jak amoniak powstałe w wyniku metabolizmu są prawdopodobnie wydalane poprzez dyfuzję do wody w odwrotnym kierunku.

6. Osmoregulacja u Paramecium Caudatum:
Paramecium Caudatum ma dwie wakuole kurczliwe, jedną przednią i jedną tylną. Funkcją wakuoli kurczliwych jest osmoregulacja, tj. regulowanie zawartości wody w organizmie i może służyć również w wydalaniu odpadów azotowych, takich jak mocznik i amoniak.
Nadmiar wody (z powodu ciągłej endosmozy) w cytoplazmie jest wydzielany do kanalików retikulum endoplazmatycznego i przechodzi do kanalików nefrydialnych → kanalików doprowadzających → i zbiera się w ampulli serii 6 do 11 promienistych kanalików, które zbiegają się w kierunku i odprowadzają do każdej wakuoli. Kanały są najbardziej widoczne podczas tworzenia się wakuoli.
Gdy każda wakuola jest obrzęk (rozkurcz) do pewnego rozmiaru, to umowy (skurcz) i odprowadza na zewnątrz prawdopodobnie przez por. Wakuole kurczliwe kurczą się naprzemiennie, w odstępach 10 – 20 sekund.
Wakuola kurczliwa tylna pracuje szybciej niż wakuola przednia z powodu pobierania dużej ilości wody do tylnej części przez cytofarynkę. Wakuole kurczliwe utrzymują optymalne stężenie wody w cytoplazmie ciała, pozbywając się jej nadmiaru.

Zachowanie Paramecium Caudatum:
Reakcje Paramecium Caudatum na różne rodzaje bodźców są poznawane przez badanie jego reakcji i grupowania się lub rozpraszania osobników w hodowli. Odpowiedź jest pozytywna, jeśli zwierzę porusza się w kierunku bodźca i negatywne, gdy porusza się z dala. Do niekorzystnego bodźca zwierzę nadal daje reakcję unikania, aż do ucieczki.
W reakcji unikania, rytm rzęskowy ulega odwróceniu, zwierzę porusza się do tyłu na krótką odległość, a następnie obraca się w kierunku stożkowym poprzez odchylenie przedniego końca do tyłu, podczas gdy obraca się na tylnej końcówce. Wszystkie regulacje dokonywane są metodą prób i błędów. Eksperymenty wykazały, że przedni koniec zwierzęcia jest bardziej wrażliwy niż inne części.
Reakcje Paramecium na różne bodźce mogą być pogrupowane w następujący sposób:
(i) Reakcje na kontakt (Thigmotaxis):
Reakcje na kontakt są zróżnicowane u Paramecium. Jeśli przedni koniec jest lekko dotknięty cienkim punktem, występuje silna reakcja unikania. Kiedy pływające Paramecium zderzy się z jakimś obiektem w wodzie, ale jeśli zostanie dotknięte w innym miejscu, może nie być żadnej reakcji. Wolno poruszający się osobnik często reaguje pozytywnie na kontakt z obiektem, zatrzymując się na nim.
(ii) Reakcje na substancje chemiczne (chemotaksja):
Generalnie Paramecia odpowiadają na bodźce chemiczne za pomocą unikania reakcji. Jeśli kropla słabego roztworu soli (0,5 procent) jest wprowadzona do populacji Paramecium na mikroprzesuwce, zwierzęta reagują reakcją unikania i żadne nie wchodzi do kropli. Natomiast w przypadku kwasów reakcja jest pozytywna, nawet jeśli stężenie jest wystarczająco silne, aby je zabić.
(iii) Reakcje na temperaturę (termotaksja):
Paramecium poszukuje optymalnej temperatury od 24 do 28°C. Kiedy zmiana temperatury następuje wyraźnie powyżej lub poniżej optymalnego zakresu, Paramecia wykazują reakcję unikania. Większe ciepło stymuluje szybki ruch i unikanie reakcji, aż do ucieczki lub zabicia zwierząt.
(iv) Reakcje na światło (fototaksja):
Z wyjątkiem zielonego Paramecium bursaria, który jest pozytywnie fototaktyczny, inne gatunki są obojętne na zwykłe światło. Jednakże, gdy natężenie światła jest nagle i gwałtownie zwiększone, na ogół następuje reakcja negatywna. Paramecia wykazują natychmiastową negatywną reakcję na promienie ultrafioletowe.
(v) Reakcje na prąd elektryczny (galwanotaksja):
Paramecia reagują na bodźce elektryczne. Kiedy dwie elektrody są umieszczone naprzeciwko siebie w płytkim naczyniu zawierającym Paramecia i przyłożony jest stały prąd, wszystkie organizmy pływają w tym samym kierunku w kierunku katody lub elektrody ujemnej, gdzie skupiają się w dużej liczbie.
Jeśli kierunek prądu elektrycznego jest odwrócony, podczas gdy Paramecia pływają w kierunku katody, organizmy odwracają kierunek i pływają w kierunku nowej katody.

(vi) Reakcje na prąd wody (reotaksja):
Paramecia wykazują dodatnią reotaksję. W łagodnym prądzie wodnym Paramecia w większości poruszają się z prądem, a ich przednie końce płyną w górę rzeki.
(vii) Reakcje na grawitację (Geotaksja):
Paramecia generalnie wykazują negatywną reakcję na grawitację, co widać w hodowli, gdzie wiele osobników gromadzi się blisko pod warstwą powierzchniową z ich przednimi końcami u góry. Jeśli Paramecia zostaną wprowadzone do odwróconej, wypełnionej wodą rurki w kształcie litery U, zatkanej na obu końcach, natychmiast przemieszczają się w górę do poziomej części rurki.
Reprodukcja u Paramecium Caudatam:
Paramecium Caudatum rozmnaża się bezpłciowo przez poprzeczne rozszczepienie binarne, a także ulega kilku typom reorganizacji jądrowej, takim jak koniugacja, endomiksja, autogamia, cytogamia i hemixis, itp.
(i) Poprzeczne rozszczepienie binarne:
Poprzeczne rozszczepienie binarne jest najczęstszym typem rozmnażania bezpłciowego u Paramecium. Jest to wyraźnie unikalny proces bezpłciowy, w którym jeden w pełni rozwinięty osobnik dzieli się na dwa osobniki bez pozostawiania zwłok rodzicielskich.
Płaszczyzna podziału jest przez środek komórki i w płaszczyźnie pod kątem prostym do długiej osi ciała. Podział ciała komórkowego jako całości jest zawsze poprzedzony podziałem jąder; rzeczywiście wydaje się, że rozmnażanie jest inicjowane przez aktywność jądrową i podział.

Paramecium caudatum rozmnaża się przez poprzeczne rozszczepienie binarne w sprzyjających warunkach. W rozszczepieniu binarnym, mikrojądro dzieli się przez mitozę na dwa mikrojądra córki, które przemieszczają się do przeciwległych końców komórki. Makronukleus wydłuża się i dzieli poprzecznie przez amitozę.
Kolejna cytofarynka ulega pączkowaniu i pojawiają się dwie nowe wakuole kurczliwe, jedna w pobliżu końca przedniego, a druga w pobliżu końca tylnego. W międzyczasie pojawia się bruzda zwężająca w pobliżu środka ciała i pogłębia się aż do całkowitego podziału cytoplazmy.
Powstałe dwie „córki” paramecia są jednakowej wielkości, każda zawiera zestaw organelli komórkowych. Z dwóch powstałych paramecji, przednia nazywana jest proter, a tylna opisthe. Rosną one do pełnego rozmiaru zanim nastąpi kolejny podział.
Proces rozszczepienia binarnego wymaga około dwóch godzin, aby zakończyć i może wystąpić jeden do czterech razy dziennie, dając 2 do 16 osobników. W ciągu roku powstaje około 600 pokoleń.
Szybkość rozmnażania zależy od warunków zewnętrznych żywności, temperatury, wieku kultury i gęstości zaludnienia, a także od wewnętrznych czynników dziedziczności i fizjologii. Naturalnie, gdyby wszyscy potomkowie jednego osobnika przeżyli i rozmnażali się, liczba wyprodukowanych paramecia szybko zrównałaby się z objętością ziemi.
Termin klon jest używany w odniesieniu do wszystkich osobników, które zostały wyprodukowane z jednego osobnika przez rozszczepienie. Wszyscy członkowie klonu są dziedzicznie tacy sami.
(ii) Koniugacja:
Ordynarnie Paramecium Caudatum rozmnaża się przez rozszczepienie binarne przez długie okresy czasu, ale w odstępach czasu może to być przerwane przez połączenie się dwóch osobników wzdłuż ich powierzchni oralnych dla seksualnego procesu koniugacji.
Koniugacja jest zdefiniowana jako tymczasowe zjednoczenie dwóch osobników, które wzajemnie wymieniają mikro materiał jądrowy. Jest to unikalny typ procesu płciowego, w którym dwa organizmy rozdzielają się wkrótce po wymianie materiału jądrowego.
Sonneborn (1947), na podstawie zachowań godowych Paramecium Caudatum, stwierdził, że każdy gatunek Paramecium istnieje w pewnej liczbie odmian lub syngenów. Ponadto, w obrębie każdego syngenu istnieje pewna liczba typów kojarzenia, zwykle dwa.
Typy godowe pozostają morfologicznie identyczne, ale wykazują różnice fizjologiczne. U P. aurelia jest 14 syngenów i 28 typów godowych, podczas gdy u P. caudatum jest 16 syngenów i 32 typy godowe. Zauważono, że zazwyczaj paramecia nie łączą się ani z przedstawicielami własnego typu godowego, ani z innymi odmianami, a jedynie z drugim typem godowym własnej odmiany.

Faktory wywołujące koniugację:
Faktory indukujące koniugację różnią się w zależności od gatunku, ale niektóre z nich są podane poniżej:
1. Koniugacja zachodzi zwykle w niekorzystnych warunkach życia; głód lub niedobór pokarmu i szczególna dieta bakteryjna lub niektóre substancje chemiczne mogą wywołać proces koniugacji u niektórych gatunków Paramecium.
2. koniugacja występuje po około 300 bezpłciowych pokoleń rozszczepienia binarnego, lub to na przemian z rozszczepienia binarnego w długich odstępach czasu, aby odmłodzić umierającego klonu, tj. występuje w osobnikach, które muszą przejść przez pożądaną liczbę bezpłciowych pokoleń, powiedział, że jest okres niedojrzałości, a następnie stają się dojrzałe płciowo do koniugacji.
3) Koniugacja zachodzi, gdy następuje zmiana w stanie fizjologicznym paramecia, wtedy zachodzi między takimi osobnikami, które mają nieco mniejsze rozmiary (210 mikronów długości) i są na etapie, który można uznać za okres niezdrowej starości; paramecia w tym stanie umrą, jeśli nie pozwoli się im na koniugację.
4) Mówi się, że nagłe zaciemnienie w warunkach oświetlenia i niskie temperatury wywołują proces koniugacji u niektórych gatunków.
5. Koniugacja nie odbywa się w nocy ani w ciemności; zaczyna się wczesnym rankiem i trwa do popołudnia.
6. Mówi się, że substancja białkowa w rzęskach osobników typu godowego wywołuje koniugację.
Proces koniugacji:
Proces koniugacji różni się u różnych gatunków Paramecium, ale niżej podana relacja dotyczy procesu koniugacji u P. caudatum (ryc. 20.21).
W koniugacji dwa Paramecium caudatum (określane jako prekoniuganty) przeciwnych typów godowych tej samej odmiany zbliżają się do siebie swoimi powierzchniami brzusznymi i łączą się rowkami gębowymi; ich rzęski wytwarzają na powierzchni ciała substancję, która powoduje przyleganie dwóch sprzęgających się paramecji.
Przestają żerować, a ich aparat gębowy zanika. W miejscu zetknięcia się obu osobników pęka otoczka i ektoplazma, a między nimi tworzy się most protoplazmatyczny. Teraz osobniki te nazywane są koniugantami.
W tym stanie sprzężona para pływa aktywnie i jednocześnie w każdym sprzężonym osobniku zachodzi szereg zmian jądrowych, opisanych poniżej:
Makronukleus zaczyna się rozpadać, staje się luźny w konsystencji i tworzy skomplikowany skręcony szkielet, w drugiej połowie okresu koniugacji w końcu zniknie będąc wchłoniętym do cytoplazmy. Mikrojądro każdego koniuganta dzieli się dwukrotnie, przy czym jeden z nich jest podziałem redukcyjnym.
W ten sposób w każdym koniugancie powstają cztery haploidalne mikrojądra córki. Trzy z tych czterech mikrojąder degenerują się w każdym z nich, tak że pozostaje tylko jedno.
Pozostałe mikrojądro każdego koniugantu dzieli się mitotycznie na dwa nierówne pronukleje lub jądra gametyczne, tworząc większe stacjonarne projądro żeńskie i mniejsze, aktywne migrujące projądro męskie.
Migrujące projądro jednego z koniugantów przekracza mostek protoplazmatyczny i łączy się ze stacjonarnym projądrem drugiego koniuganta, tworząc synkarion lub jądro koniugacyjne, w którym przywrócona jest diploidalna liczba chromosomów i nastąpiła wymiana materiału dziedzicznego.
Proces ten został porównany z zapłodnieniem u zwierząt wyższych, ale nie jest to zapłodnienie, ponieważ nie ma tu gamet. Koniuganty teraz (po około 12-48 godzinach) rozdzielają się i są nazywane eks-koniugantami. Synkarion każdego byłego koniuganta dzieli się trzykrotnie, tworząc osiem mikrojąder w każdym byłym koniugancie.
Cztery z ośmiu mikrojąderek powiększają się i stają się makrojądrami, a trzy z pozostałych czterech mikrojąderek znikają.
Pozostałe mikrojądro dzieli się i w tym samym czasie eks-koniugant dzieli się przez rozszczepienie binarne na dwie komórki, z których każda ma dwa makrojądra i jedno mikrojądro. Komórki i ich mikrojądra dzielą się po raz drugi, tworząc cztery paramecia z każdego byłego koniuganta, tak że każda ma jedno makrojądro i jedno mikrojądro.
Nowe makrojądro, jak również mikrojądro, zostały wykonane z nowego materiału. Te nowe jądra zawierają prawdopodobnie nowy i inny potencjał, który znajduje odzwierciedlenie u zdrowych osobników.
Znaczenie koniugacji:
Klon wymrze, jeśli nie nastąpi reorganizacja jądrowa, ale klon można odmłodzić, aby odzyskał dawny wigor przez rearanżację jądrową, tę reorganizację jądrową przynosi koniugacja, a więc koniugacja jest niezbędna dla kontynuacji rozszczepienia binarnego.
Znaczenie koniugacji została podsumowana poniżej:
1. Koniugacja służy jako proces odmładzania i reorganizacji, dzięki któremu przywracana jest witalność rasy. Jeśli koniugacja nie zachodzi przez dłuższy czas, paramecia słabną i umierają. (Twierdzenie Woodruffa o utrzymywaniu paramecia w zdrowiu przez 22,000 pokoleń bez koniugacji zostało obalone przez Sonneborna, ponieważ wykazał on, że wszystkie paramecia Woodruffa należały do tego samego typu kojarzenia).
2. Nie ma rozróżnienia płci u koniugantów, chociaż tylko paramecia z dwóch różnych typów godowych tej samej odmiany będą się koniugować.
3. Nie ma rozróżnienia płci, ale aktywne pro-jądro wędrowne jest uważane za męskie, a pro-jądro stacjonarne za żeńskie.
4. koniugacja jest tylko tymczasowym zjednoczeniem, nie ma fuzji cytoplazmy i nie powstaje zygota, ale jądro każdego byłego koniuganta zawiera materiał dziedziczny pochodzący od dwóch koniugujących osobników.
5. Koniugacja powoduje zastąpienie makrojądra materiałem pochodzącym z synkarionu, jest to wydarzenie o podstawowym znaczeniu. W rozszczepieniu binarnym chromosomy makrojądra były rozdzielane losowo do komórek potomnych, dalsze rozszczepienie binarne powodowało, że klon był słaby, z pewnymi nieprawidłowościami strukturalnymi.
Konjugacja przynosi tworzenie prawidłowej liczby chromosomów w makrojądrze, tak, że wyścig jest odnowiony w wigorze. Rolą mikrojądra jest przywrócenie równowagi kompleksu chromosomów i genów.
Zachowanie jądrowe w reprodukcji u Paramecium Caudatum:
Paramecium Caudatum wykazuje pewne odchylenia w zachowaniu jądrowym podczas rozszczepienia i koniugacji, tymi odchyleniami są endomiksja, autogamia, cytogamia i hemixis. W pierwszych trzech procesach dochodzi do rekombinacji genetycznej i z mikrojądra powstaje nowe makrojądro.
(i) Endomiksja:
Woodruff i Erdmann (1914) przede wszystkim donieśli o nowym procesie reorganizacji jądrowej, endomiksis (gr., endon = wewnątrz; mixis = mieszanie) u Paramecium aurelia, gatunku dwujądrzastego (ryc. 20.22). Proces ten został opisany jako występujący okresowo, w którym nowy aparat makrojądrowy jest wytwarzany bez tworzenia synkarionów. Endomiksja występuje w obrębie jednego osobnika.
Według Woodruffa i Erdmanna makronukleus degeneruje się, a mikrojądra dzielą się dwukrotnie, tworząc osiem mikrojąder. Sześć mikronuklei degeneruje i dwa pozostają. Gdy pozostają tylko dwa mikrojądra, zwierzę dzieli się przez rozszczepienie na dwie komórki, każda z jednym mikrojądrem.
Mikronukleus każdej komórki córki dzieli się dwukrotnie, tworząc cztery mikronuklei. Dwa mikrojądra powiększają się tworząc makronukleje. Zwierzę i jego mikrojądra dzielą się tak, że powstają dwa osobniki-córki, z których każdy ma jedno makrojądro i dwa mikrojądra.
Endomixis występuje w tej odmianie P. aurelia, która nie koniuguje, stąd efekt endomixis może być taki sam jak koniugacji, ponieważ oba procesy przynoszą zastąpienie makrojądra materiałem z mikrojądra, i oba procesy odmładzają żywotność rasy.
Ale te dwa procesy różnią się, ponieważ w endomiksie nie ma fuzji pronuklei; endomiksja może być porównana do partenogenezy.
Jednakże niektórzy pracownicy twierdzą z uzasadnionych powodów, że endomiksis nie jest ważnym procesem i została opisana na podstawie błędnych obserwacji. Najprawdopodobniej endomiksis nie zachodzi i może być tylko wyspecjalizowanym przypadkiem autogamii.

Później, Erdmamm i Woodruff (1916) donieśli o endomiksis u Paramecium caudatum. Diller, jednakże, nie wierzy w ważność tego procesu i uważa, że Erdmann i Woodruff po prostu połączyli etapy hemixis i autogamii w jeden schemat, endomixis.
(ii) Autogamia:
Diller (1934, 1936) i Sonneborn (1950) opisali proces samozapłodnienia lub autogamii zachodzący u pojedynczego osobnika u Paramecium aurelia (ryc. 20.23). Podał on, że w autogamii trzy podziały mikrojądrowe (pregamiczne), obejmujące dojrzewanie, powodują powstanie jąder gametycznych (pronuklei).
Podczas autogamii u P. aurelia dwa mikrojądra dzielą się dwukrotnie (raz mejotycznie), tworząc osiem mikrojąder, z których sześć ulega degeneracji. W międzyczasie makrojądro rozrasta się w masę przypominającą szkielet, która pęka na kawałki i zostaje wchłonięta do cytoplazmy. Dwa z ośmiu mikrojąder, jako pronukleje, wchodzą do stożka protoplazmatycznego wybrzuszającego się w pobliżu ujścia komórki.
Dwa pronukleje łączą się, tworząc synkarion. Synkarion dzieli się dwukrotnie, tworząc cztery mikrojądra. Dwa mikrojądra stają się makrojądrami. Paramecium i jego mikrojądra dzielą się tworząc dwa osobniki potomne, każdy z jednym makrojądrem i dwoma mikrojądrami. Proces ten jest zakończony w ciągu około dwóch dni.
Autogamia powoduje odmłodzenie rasy.
Przypomina ona koniugację o tyle, że nowe makrojądro powstaje z materiału pochodzącego z mikrojądra, w nowym makrojądrze zostaje przywrócona prawidłowa liczba chromosomów; a także w tym, że następuje fuzja dwóch pronuklei. Ale autogamia różni się od koniugacji tym, że w autogamii bierze udział tylko jeden osobnik, który dostarcza obu pronukleusów, jest to rodzaj samozapłodnienia.
(iii) Hemixis:
Diller (1936) podał występowanie hemixis u Paramecium aurelia (ryc. 20.24). Hemiksis jest przede wszystkim procesem fragmentacji i podziału makrojądrowego bez żadnej niezwykłej aktywności mikrojądrowej. Diller sklasyfikował hemixis na cztery typy, mianowicie A, B, C i D, jak pokazano na Rys. 20.24 u P. aurelia, ale napotkał również wszystkie typy w masowych hodowlach P. caudatum i P. multimicronucleatum.
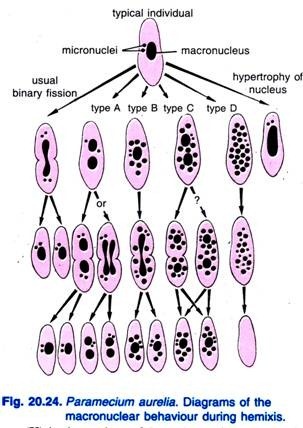
Typ A jest najprostszą formą hemixis charakteryzującą się podziałem makronukleusa na dwie lub więcej części. Podział ten nie jest zsynchronizowany z podziałem mikrojądrowym.
Typ B charakteryzuje się wyciskaniem od jednej do 20 lub więcej kulek chromatyny z makrojądra do cytoplazmy.
Typ C charakteryzuje się jednoczesnym rozszczepieniem makrojądra na dwie lub więcej głównych części i wyciskaniem kulek makrojądrowych do cytoplazmy.
Typ D jest uważany za reprezentujący warunki patologiczne, w których makrojądro ulega całkowitej fragmentacji na kulki chromatyny, które ostatecznie znikają z komórki. Mikrojądra na ogół znikają przed rozpuszczeniem makrojądra.
(iv) Cytogamia:
Wichterman (1939) doniósł o innym procesie płciowym u Paramecium caudatum, który nazwał cytogamią. W cytogamii, nie ma wymiany jądrowej. W tym procesie dwa osobniki zbliżają się do siebie powierzchniami brzusznymi, ale otoczka obu osobników nie ulega rozpadowi.
Mikronukleus każdego osobnika dzieli się trzykrotnie, tworząc osiem mikrojąder, z których sześć rozpada się u każdego osobnika. Dwa pozostałe mikrojądra łączą się, tworząc synkarion w każdej komórce. Zwierzęta teraz się rozdzielają.
Cytogamia różni się od autogamii w tym, że są dwa zwierzęta w kontakcie ze sobą, ale przypomina autogamię i koniugacji w fuzji dwóch pronuclei. Cytogamia różni się od koniugacji tym, że nie ma wymiany jądrowej między dwoma zwierzętami, które się spotykają.
Niektóre cząsteczki cytoplazmatyczne zgłoszone w Paramecium Caudatum:
(i) Cząsteczki Kappa:
W 1938 roku T.M. Sonneborn poinformował, że niektóre rasy (znane jako zabójcy lub szczep zabójcy) Paramecium wytwarzają trującą substancję, zwaną paramecyną, która jest śmiertelna dla innych osobników zwanych wrażliwcami. Paramecyna jest rozpuszczalna w wodzie, dyfuzyjna i zależy od jej produkcji na niektórych cząsteczkach znajdujących się w cytoplazmie Paramecium (szczep zabójcy).
Cząsteczki te nazywane są cząsteczkami kappa. Cząsteczki kappa mają DNA i RNA. Zabójczy Paramecium może zawierać setki cząstek kappa. Szczegółowe badania tych cząsteczek ujawniły, że dominujący gen (K) w jądrze Paramecium jest niezbędny, aby cząsteczki kappa mogły istnieć, rozmnażać się i produkować paramecynę.
(ii) mµ, cząsteczki:
R.W. Siegel (1952) doniósł o innym typie zabójczych cząstek w cytoplazmie niektórych Paramecium. Paramecium z cząsteczkami mp jest nazywane mate killer, ponieważ kiedy łączy się z Paramecium bez cząsteczek mµ zwanych mate sensitive, zabija to ostatnie. Cząsteczki mp składają się również z DNA, RNA, itd.
Cząsteczki te istnieją tylko w tych parameciach, których mikrojądro zawiera co najmniej jeden gen dominujący jednej z dwóch par niepowiązanych genów chromosomalnych (M1 i M2).
(iii) Cząsteczki Pi:
Cząstki te mają być zmutowaną formą cząstek kappa, ale nie wytwarzają żadnego rodzaju trującej substancji.
(iv) Cząstki lambda:
Cząsteczki te są zgłaszane w parameciach zabójczych i mówi się, że produkują jakąś substancję odpowiedzialną za powodowanie lizy lub rozpadu paramecji wrażliwych, tj. takich, które jej nie posiadają.