- Volvox Case Study Guide Covers up Classification, Characteristics, Volvox Under Microscope, Reproduction (Asexual and Sexual Reproduction)
- Detailed Guide With All Aspects
- Volvox Klasyfikacja
- Volvox Charakterystyka
- Ciało roślinne (gametofit)
- Struktura komórki
- Volvox Unicellular lub Multicellular
- Volvox Under Microscope
- Volvox Reprodukcja
- Rozród bezpłciowy
- Rozród płciowy
Volvox Case Study Guide Covers up Classification, Characteristics, Volvox Under Microscope, Reproduction (Asexual and Sexual Reproduction)
Volvox jest słodkowodną planktoniczną (swobodnie pływającą) algą. Istnieje około 20 gatunków należących do tych rodzajów. W królestwie roślin występuje jako najpiękniejszy i najbardziej atrakcyjny obiekt. Występuje w słodkiej wodzie jako zielone kulki wielkości główki szpilki. Są one widoczne gołym okiem, około 0,5 mm. in diameter.
Detailed Guide With All Aspects
Alga ze względu na specyficzny wzór pływania jest często określana jako, rolling alga. Jej wzrost jest często obserwowany w tymczasowych lub stałych stawach słodkowodnych, basenach, rowach, a także w jeziorach. Wzrost jest obfity, gdy temperatura i materia organiczna jest dostępna w wystarczającej ilości. Jego rozmnażanie jest tak częste i obfite, że woda w stawach staje się zielona (zakwit wody). Wiosna i pory deszczowe są typowymi okresami jego aktywnego wzrostu wegetatywnego. Z nadejściem niekorzystnego okresu (lato) alga zanika i przechodzi niekorzystny okres w postaci zygoty. Volvox zrobić ich własne jedzenie przez fotosyntezę.
Volvox Klasyfikacja
- Klasa: Chlorophyceae
- Order: Volvocales
- Podrząd: Chlamydomonadineae
- Rodzina: Sphaerellaceae
- Genus: Volvox
Najczęściej występującymi gatunkami Volvox są V, globator, V, aureus; V. prolificus, V. rouseletti, V. merelli.
Volvox Charakterystyka
Ciało roślinne (gametofit)
Volvox jest koenobialnym zielonym glonem, {(ciało kolonijne-roślinne nie ma stałej liczby komórek np. Volvox aureus) (ciało koenobialne-roślinne mające stałą liczbę komórek, np, Pandorina moruma, liczba komórek wynosi 4, 8, 16 lub 32. Eudorina unicocca, liczba komórek 16, 32 lub 64)}. Wśród form ruchliwych koenobium Volvoxa jest największą, wysoce zróżnicowaną i dobrze rozwiniętą algą. Każde coenobium jest elipsoidalnym lub pustym w środku ciałem kulistym z dokładnie zaznaczoną delikatną śluzowatą warstwą definitywną. Wewnętrzna część coenobium składa się z dyfuzyjnego (wodnistego) śluzu, podczas gdy komórki ułożone są w pojedynczą warstwę na peryferiach.
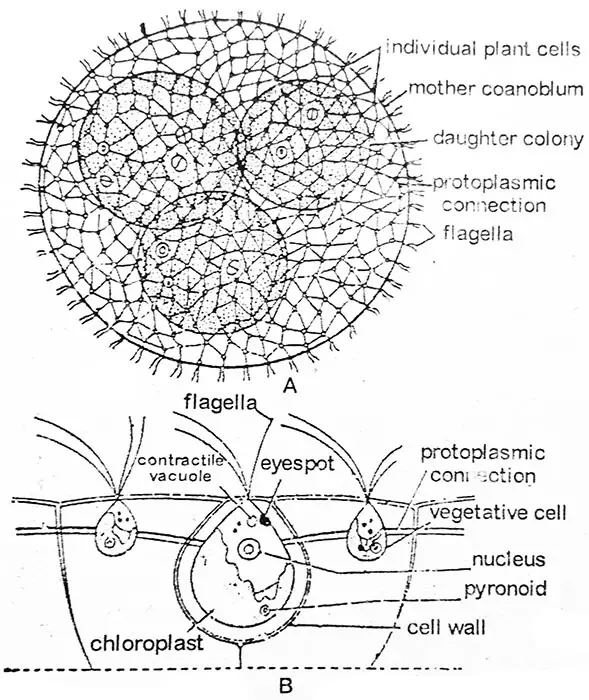
Ruch kolonii wywołany jest wspólnym działaniem flagelli poszczególnych komórek. Każde coenobium ma określony koniec przedni i tylny. Coenobium wykazuje biegunowość, porusza się i obraca powoli, wykazując niezwykłą współpracę między komórkami przedniego i tylnego końca w trakcie swojego ruchu. Volvox nie jest osobnikiem, ale zrzeszeniem wielu podobnych komórek, z których każda funkcjonuje jak niezależny osobnik i prowadzi własne odżywianie, oddychanie i wydalanie i nie wykazuje współpracy między komórkami w tych funkcjach. Liczba komórek na coenobium waha się np. 500-1000 u V. aureus, 1500-20,000 u V. globator i nawet do około 60,000 u V. rouseletti.
Struktura komórki
Według badań każda pojedyncza komórka ma swoją tożsamość. Jest on otoczony przez własne duże galaretowate, osłony, które mogą być sprzeczne z osłonami sąsiednich komórek lub mogą być odrębne od siebie. Wówczas są one odrębne, są one kanciaste przez wzajemne uciskanie i mają zwykle zarys sześciokątny. Rys.,2.22. W ten sposób znaczna przestrzeń galaretowatego materiału pomaga w oddzieleniu jednej komórki od drugiej. U większości gatunków każda komórka jest połączona z komórkami sąsiednimi szeregiem nitek protoplazmatycznych lub cytoplazmatycznych (plasmodesmae Rys. 2.22) powstałych w trakcie podziałów komórkowych i rozwoju kolonii. Nitki protoplazmatyczne mogą być cienkie i delikatne u V. aureus, okrągłe u V. globator; lub klinowate u V. mononae, lub mogą być nieobecne, jak u V. tertius.
W zarysie pojedyncza komórka Volvox przypomina Chlamydomopas. Każda komórka ma przednio włożone parę flagelli o równej długości. Obie flagelle są typu whiplash. Flagi wystają poza powierzchnię koenobium do otaczającej je wody. W pobliżu podstawy flagelli znajdują się dwie lub więcej wakuol kurczliwych. Protoplast ma różne kształty w zależności od gatunku. W V. tertius protoplasta w V. aureus jest zaokrąglony i typu Chlamydomonas, podczas gdy w V. globator protoplasta jest gwiaździsty typ o rozproszonych chloroplastów i rozproszonych wakuoli kurczliwych.
Volvox Unicellular lub Multicellular
Wegetatywne komórki młodej kolonii są zielone i podobne w wielkości i kształcie, ale w starszych koloniach, niektóre komórki tylnej części regionu wzrost dziesięć razy; lub więcej wielkości normalnej komórki. Rozwijają one liczne pyrenoidy zwiększają swój rozmiar i zachowują się jak komórki rozrodcze. Mogą być bezpłciowe lub, płciowe. W niektórych przypadkach w tej samej kolonii mogą występować zarówno komórki bezpłciowe jak i płciowe. W części przedniej komórki kolonii pozostają tylko wegetatywne i są stosunkowo mniejsze. W części przedniej komórki noszą większą plamkę oczną. Tak więc kolonia składa się z dwóch typów komórek: komórek rozrodczych i komórek somatycznych.
Volvox może służyć jako przykład pierwszego kroku w kierunku koordynacji i podziału pracy. Kolonia Volvox może być traktowana jako typ wielokomórkowy składający się z komórek wyodrębnionych do wykonywania różnych funkcji. Komórki pełniące różne funkcje to,
- Komórki wegetatywne zajmujące się wytwarzaniem pokarmu i biorące udział w lokomocji,
- Komórki bezpłciowe produkujące kolonie córki
- Komórki płciowe: produkujące jaja, oraz produkujące pylniki
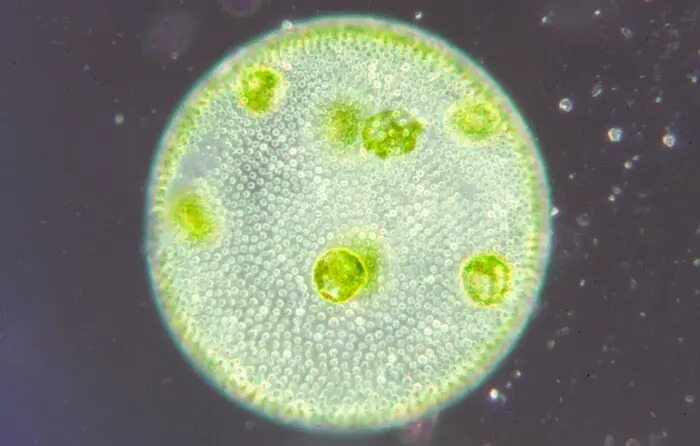
Volvox Under Microscope
Poniżej przedstawiono widok mikroskopowy kolonii volvox:
Volvox Reprodukcja
W przeciwieństwie do Chlamydomonas, komórki kolonii Volvox wykazują specjalizację funkcjonalną. Rozmnaża się ona zarówno bezpłciowo jak i płciowo. Na początku sezonu wegetacyjnego (sprzyjające warunki). rozmnażanie jest bezpłciowe. To właśnie z tego powodu wszystkie kolonie zebrane w jednym czasie są albo bezpłciowe albo płciowe.
Rozród bezpłciowy
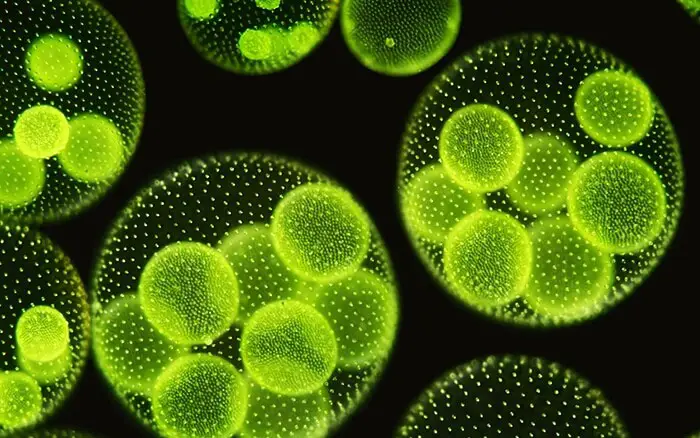
Rozród bezpłciowy występuje na początku sezonu wegetacyjnego podczas sprzyjających warunków. We wcześniejszych etapach, wszystkie komórki kolonii są podobne, ale później, kilka komórek w tylnej połowie kolonii przechowywać żywności i wzrost w rozmiarze. Te znacznie powiększone komórki są wyspecjalizowanymi komórkami bezpłciowymi zwanymi gonidiami (w liczbie pojedynczej gonidium). Ich liczba waha się od dwóch do pięćdziesięciu w pojedynczym coenobium.
Rozwój córki coenobium z gonidium
Przed podziałem, gonidia są lekko wepchnięte do wnętrza kolonii i można je odróżnić jako rząd komórek wegetatywnych dzięki większym rozmiarom, zaokrąglonemu kształtowi, braku flagelli i plamki ocznej, wyraźnemu jądru, kilku pyrenoidom i gęstej ziarnistej cytoplazmie. Każde gonidium leży w kulistej, galaretowatej osłonce.
Pierwszy podział protoplastu gonidialnego jest podłużny, tzn. przebiega w płaszczyźnie przednio-tylnej coenobium. Drugi podział jest również podłużny i przebiega pod kątem prostym do pierwszego. Każda z powstałych w ten sposób komórek potomnych dzieli się ponownie podłużnie, tak że powstaje płytka ośmiokomórkowa. Jest ona zakrzywiona, a jej wklęsła powierzchnia skierowana jest na zewnątrz. To ośmiokomórkowe stadium znane jest jako stadium Plakea. Równoczesne podłużne podziały komórek potomnych trwają przez kilka pokoleń komórek (do 14, 15 lub 16 razy u V. rouseletti).
Na szesnastokomórkowym etapie, komórki są ułożone na obwodzie pustej kuli, z małym otworem, phialopore w kierunku zewnętrznym macierzystego coenobium. Na tym etapie wszystkie komórki są nagie i stykają się ze sobą. Ich przednie końce zwrócone są w kierunku centrum sfery. Wraz z postępem inwazji fialopory znacznie się powiększają. Jak in-folding tylnej części (invagination) zaczyna przepychać się przez phialopore, jego otaczające krawędzie są zawinięte do tyłu, które stopniowo przesuwają się w dół, aż cała struktura jest odwrócony. Fiolopor, który teraz wykazuje szereg fałd, stopniowo staje się zamknięty. Proces inwersji wymaga około trzech do pięciu godzin. W niektórych nienormalnych przypadkach inwersja w ogóle nie zachodzi, jak odnotowano u V. minor.
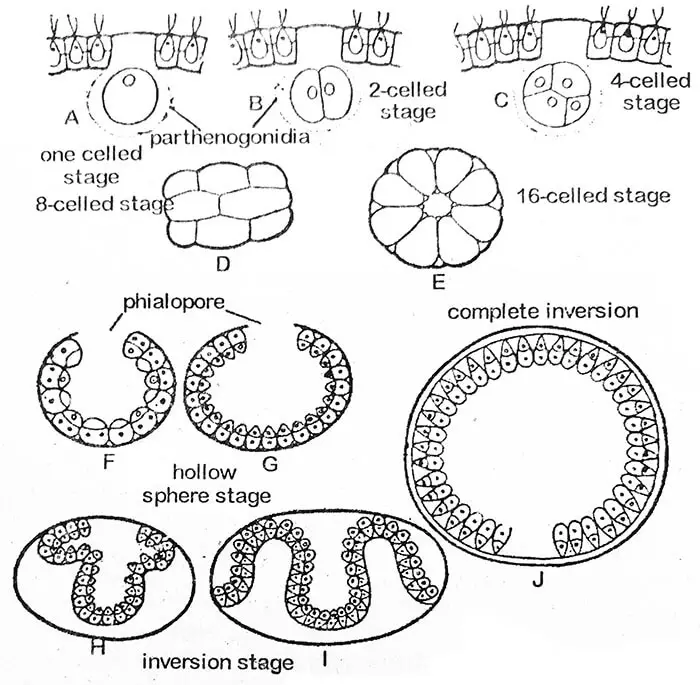
Komórki córek coenobium zaczynają się teraz oddzielać od siebie poprzez rozwój śluzowatych części (ściana komórkowa). Każda komórka nabywa w końcu parę flagelli i błonę komórkową. Kolonia potomna (coenobium) jest nadal utrzymywana w obrębie macierzystej ściany komórkowej, która ostatecznie przekształca się w otaczającą ją śluzowatą błonę. W jednej kolonii macierzystej może rozwijać się jednocześnie kilka kolonii potomnych. W ten sposób mogą one wypełniać pusty, środkowy obszar kolonii macierzystej.
Kolonie córek są uwalniane wraz ze śmiercią i rozpadem kolonii macierzystej. Czasami kolonie córek nie są uwalniane przez dłuższy czas i rozwijają się kolonie wnuczek. Tak więc, aż 2-4 pokolenia uwięzionych kolonii córek mogą być widoczne w jednej oryginalnej kolonii rodzicielskiej, zwłaszcza u V. africanus.
Rozród płciowy
Volvox wykazuje zaawansowany oogamiczny typ rozmnażania płciowego, który odbywa się poprzez tworzenie pylników i oogonii. Mogą one powstawać na tym samym coenobium (jednopienne) jak u V. globator lub na różnych coenobium (dwupienne) jak u V. aureus. Gatunki jednopienne są protandrous (pylniki rozwijają się jako pierwsze), dlatego u takich gatunków zapłodnienie nastąpi między pylnikiem a komórką jajową różnych roślin. Interesujące jest to, że kolonie płciowe są często pozbawione bezpłciowych kolonii córek. U koenobium komórki przeznaczone do utworzenia organów płciowych znajdują się w tylnej połowie. Są one uważane za komórki wyspecjalizowane. Narządy płciowe (gametangia) są produkowane w mniejszej liczbie. Podczas tworzenia się gametangiów komórka ulega powiększeniu i zaokrągleniu oraz pozbywa się flagelli, ale pozostaje połączona z innymi komórkami drobnymi nićmi protoplazmatycznymi. Męskie gametangtum nazywane jest antheridium, podczas gdy żeńskie oogonium.
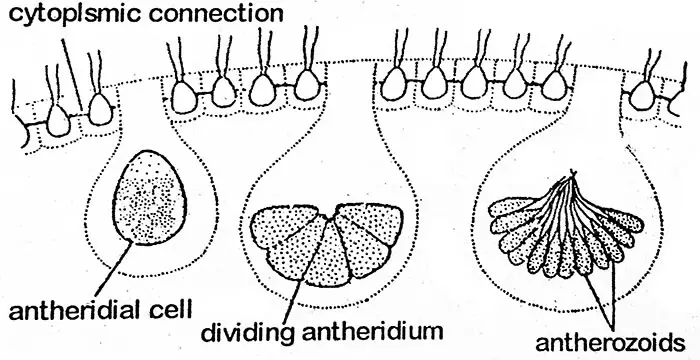
Protoplast antheridium ulega powtarzającym się podziałom komórkowym w sposób podobny do tego, jaki obserwuje się w rozwoju bezpłciowej komórki gonidialnej w kolonię córkę (tj. stadium plakea). W ten sposób powstaje masa małych, nagich, biflagellowatych, bruzdkowanych pylników w liczbie od 16 do 512 w pylniku. Są one zgrupowane jako płaska płytka, z wyjątkiem V. aureus, u którego pylniki występują w kolonii bezpłciowej. Płytka pylników ulega rozpadowi i uwalnia pylniki.
Pylnik
Każdy pylnik to dwupłatkowa, wydłużona, stożkowata lub wklęsła struktura z pojedynczym jądrem i małym żółto-zielonym lub bladozielonym chloroplastem.
Oogonium
Komórka oogonialna znacznie się powiększa i pozbywa się flagelli oraz połączeń protoplazmatycznych z sąsiednimi komórkami. Komórka staje się zaokrąglona lub w kształcie kolby, z dużą częścią wystającą do wnętrza coenobium. Na tym etapie nazywana jest oogonium, którego cała część przekształca się w pojedyncze kuliste jajo z dzióbkowatym występem po jednej stronie. Przez ten koniec do oogonium wchodzi pylnik. Jajo (oosfera) zawiera duże, centralnie umieszczone jądro i ciemieniowy chloroplast z pyrenoidami. Jest ono obficie zaopatrzone w substancje zapasowe często wchłaniane z sąsiednich komórek przez nitki protoplazmatyczne.
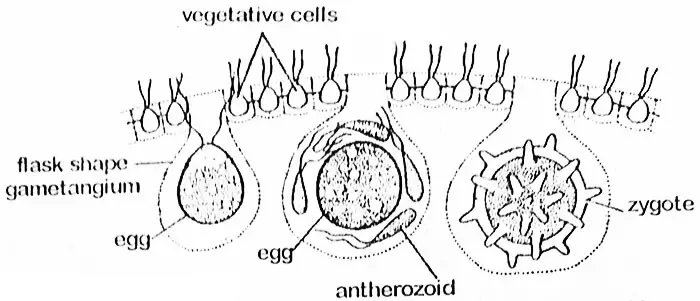
Płodnienie
Pylniki są uwalniane w grupach w czasie zapłodnienia i pozostają one nienaruszone aż do momentu dotarcia do jaja. Pylniki są wtedy uwolnione. Tylko jeden pylnik łączy się z jajkiem i powoduje powstanie oospory. Oospora wydziela następnie trójwarstwową gładką lub kolczastą ścianę. Gromadzi wystarczającą ilość hematochromu (granulki czerwonego barwnika, prawdopodobnie ksantofilu), który nadaje mu pomarańczowy wygląd. Na tym etapie może być nazywany zygotą.
Oospora i jej kiełkowanie
Zewnętrzna ściana, egzospora może być gładka, (V. globator) lub kolczasta (V. speematospaera). Warstwa środkowa to mezospory, a wewnętrzna to endospory. Zygota zawiera wystarczającą ilość zapasowego materiału pokarmowego i innych wtrąceń. Zygota jest więc zatrzymywana przez koenobium, które może być uwolnione przez rozpad galaretowatej matrycy. Po uwolnieniu osiada ona na dnie basenu i może pozostać żywotna przez kilka lat.

Na początku sprzyjających warunków zygota rozwija się w różny sposób. U V. campensis jądro zygoty dzieli się mejotycznie i tworzy cztery jądra, z których trzy degenerują się, a jedno przeżywa: Przetrwałe jądro wraz z zawartością cytoplazmatyczną wydostaje się z pęcherzyka. Na tym etapie można go nazwać rojnikiem (duża liczba lub zwarta grupa, owadów, komórek itp.) pływa on swobodnie i dzieli się i dzieli ponownie tworząc nowe coenobium. Podczas kiełkowania zewnętrzne dwie warstwy ściany stają się galaretowate, a wewnętrzna warstwa tworzy pęcherzyk, który później wypełnia się protoplastą zygoty. U V. rouseletti i V. minor protoplasta zygoty przekształca się w pojedynczą zoosporę, która w wyniku dalszych podziałów tworzy nowe coenobium. Takie coenobium składa się z mniejszej liczby komórek, które rozmnażają się bezpłciowo przez następne sześć lub więcej pokoleń, za każdym razem zwiększając liczbę w kolejnych pokoleniach.
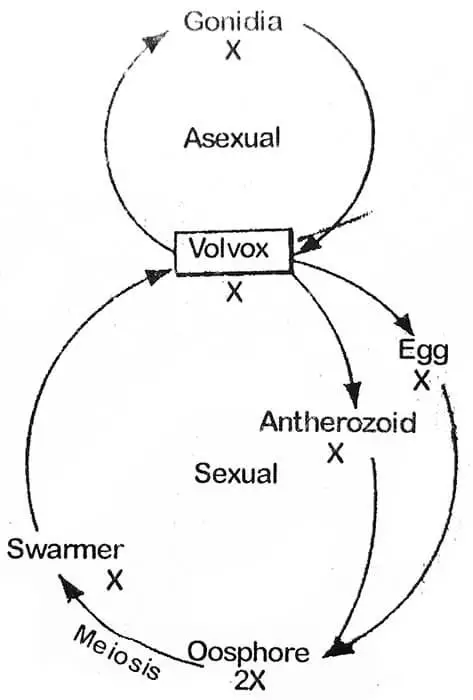
Zygota jest jedyną diploidalną fazą w cyklu życiowym Volvoxa i dlatego główny organizm rośliny jest haploidalny. Dlatego zygota musi przejść podział redukcyjny podczas formowania nowej kolonii.
.