ADVERTISEMENTS:
Neste artigo vamos discutir sobre Paramecium Caudatum:- 1. Habitat, Habitat e Cultura do Paramecium Caudatum 2. Estrutura do Paramecium Caudatum 3. Locomoção 4. Nutrição 5. Respiração e Excreção 6. Osmoregulação 7. Comportamento 8. Reprodução 9. Comportamento Aberto na Reprodução 10. Algumas Partículas Citoplasmáticas.
Conteúdo:
- Habit, Habitat e Cultura do Paramecium Caudatum
- Estrutura do Paramecium Caudatum
- Locomoção do Paramecium Caudatum
- Nutrição do Paramecium Caudatum
- Respiração e Excreção do Paramecium Caudatum
- Osmoregulação do Paramecium Caudatum Paramecium Caudatum
- Comportamento do Paramecium Caudatum
- Reprodução no Paramecium Caudatum
- Comportamento da reprodução no Paramecium Caudatum
- Partes citoplasmáticas relatadas no Paramecium Caudatum
- Habit, Habitat e Cultura do Paramecium Caudatum:
- Cultura do Paramecium:
- Estrutura do Paramecium Caudatum:
- (i) Tamanho e Forma do Paramecium Caudatum:
- (ii) Película:
- (iii) Cilia:
- (iv) Sistema Infra-Cilíndrico:
- (v) Sulco Oral e Cytopyge:
- (vi) Citoplasma:
- (vii) Ectoplasma:
- (viii) Tricocistos:
- (ix) Neuromotorium and Associated Fibrils:
- (x) Endoplasm:
- (xi) Vacúolos contráteis:
- (xii) Aspúolos para alimentos:
- Locomotion of Paramecium Caudatum:
- (i) Metabolia ou Contorções Corporais:
- (ii) Locomoção Ciliar:
- Nutrição do Paramecium Caudatum:
- Mecanismo de alimentação:
- Digestão e ingestão de Paramecium Caudatum:
- Respiração e Excreção de Paramecium Caudatum:
- Comportamento do Paramecium Caudatum:
- (i) Reacções ao contacto (Thigmotaxis):
- (ii) Reações a produtos químicos (Quimiotaxis):
- (iii) Reações à temperatura (Termotaxis):
- (iv) Reacções à luz (Fototaxis):
- (v) Reações à corrente elétrica (Galvanotaxis):
- (vi) Reações à corrente de água (Reotaxis):
- (vii) Reacções à gravidade (Geotaxis):
- Reprodução em Paramecium Caudatam:
- (i) Fissão Binária Transversa:
- (ii) Conjugação:
- Comportamento Aberrante na Reprodução em Paramecium Caudatum:
- (i) Endomixis:
- (ii) Autogamia:
- (iii) Hemixis:
- (iv) Citogamia:
- Particulas Citoplasmáticas Relatadas em Paramecium Caudatum:
- (i) Kappa Particles:
- (ii) mµ, Partículas:
- (iii) Partículas de Pi:
- (iv) Partículas de Lambda:
Habit, Habitat e Cultura do Paramecium Caudatum:
Paramecium caudatum (Gr., paramekes = oblongo; L., caudata = cauda) é comumente encontrada em lagoas de água doce, poças, valas, riachos, lagos, reservatórios e rios. É especialmente encontrada em abundância em lagoas estagnadas ricas em matéria em decomposição, em infusões orgânicas e na água de esgoto. O Paramecium caudatum é um organismo de vida livre e esta espécie está em distribuição mundial.
Cultura do Paramecium:
ADVERTISEMENTOS:
Tomar ervas daninhas submersas de um lago e colocá-las num frasco de água destilada, cobrir o frasco e deixá-lo a apodrecer; enxames de Paramecium caudatum aparecerão em poucos dias. Agora ferva o feno na água, decante a infusão e adicione alguns grãos de trigo, e deixe-o ficar até ficar turvo com bactérias.
Transfira Paramecia do primeiro frasco para este líquido, onde se multiplicarão rapidamente. Somente as infusões de feno produzirão Paramecia mostrando a presença de cistos, e cistos parecidos com grãos de areia foram relatados, mas não há provas de Paramecia formando cistos, já que eles nunca foram confirmados.
Estrutura do Paramecium Caudatum:
(i) Tamanho e Forma do Paramecium Caudatum:
O Paramecium caudatum (Fig. 20.1) é um organismo microscópico e visível a olho nu como um corpo minúsculo e alongado.
ADVERTISEMENTOS:
Parece cinza claro ou branco medindo normalmente entre 170 a 290 microns de comprimento e pode atingir um comprimento de até 300-350 microns. P. caudatum parece a sola de um chinelo ou sapato, por isso, o animal é comumente conhecido como animal-patola. É quatro vezes mais longo do que largo e um pouco cilíndrico, com extremidades nitidamente diferentes.
A parte anterior que se move para a frente é esguia com uma extremidade romba ou arredondada, enquanto que a extremidade posterior é um pouco pontiaguda ou em forma de cone. A parte mais larga do organismo está logo abaixo do meio. O corpo do animal é assimétrico em forma mostrando uma superfície oral ou ventral bem definida e uma superfície aboral ou dorsal.
(ii) Película:
O corpo é coberto por uma película fina, dupla camada, elástica e firme feita de gelatina. A película mantém a forma do animal, mas é suficientemente elástica para permitir contracções. A película tem membrana dupla, a membrana externa é contínua com a cílios e a membrana interna com o ectoplasma. Sob a maior ampliação do microscópio, a película apresenta depressões rectangulares ou hexagonais na sua superfície.

Este arranjo é preservado na superfície dorsal do Paramecium mas na superfície ventral as cristas convergem na frente e atrás em direcção a uma abertura pré e pósoral. Cada depressão hexagonal é perfurada por uma abertura central através da qual emerge um único cório. As margens anterior e posterior das depressões hexagonais suportam as aberturas dos tricocistos.
O estudo microscópico eletrônico da película (Fig. 20.2) por Ehret e Powers (1957) revelou que as depressões hexagonais correspondem a séries regulares de cavidades, os alvéolos. Todos os alvéolos formam colectivamente uma camada alveolar contínua, que é delimitada por uma membrana alveolar externa e uma alveolar interna.
A camada externa encontra-se em estreito contacto sob a membrana celular externa. Portanto, a película inclui a membrana celular externa, membrana alveolar externa e a membrana alveolar interna.
(iii) Cilia:
O corpo inteiro está coberto de numerosos, pequenos, cabelos como projecções chamadas cílios. Cílios ocorrem em filas longitudinais por todo o corpo, esta condição é conhecida como holotrichosa, na qual os cílios do corpo são iguais. Os cílios têm a mesma estrutura que os flagelos, eles têm uma bainha protoplasmática externa ou membrana plasmática com nove fibrilas longitudinais duplas em um anel periférico. Em alguns cílios, as nove fibrilas externas não são emparelhadas.
ADVERTISEMENTOS:
Existem duas fibrilas longitudinais centrais que são mais finas que as fibrilas externas. Cada cório surge de um grânulo basal ou cinetosoma. Os nove pares de fibrilas periféricas fundem-se para formar a parede do cinetosoma, assim, o cinetosoma é um tubo que está aberto ou fechado na sua extremidade inferior, as duas fibrilas centrais param ao nível da película na maioria dos ciliados.
Artindo do cinetosoma é um rizoplasto fino que não une o núcleo. Muitos Metazoários também têm cílios, a sua estrutura é a mesma, excepto que o grânulo basal é diferente e tem filamentos finos ou fibras de enraizamento que se estendem até ao citoplasma. Mas os cílios diferem dos flagelos por serem geralmente mais numerosos e de menor tamanho.
A ciliatura pode ser convenientemente dividida em corpo ou cílios somáticos que são encontrados na superfície do corpo, e em ciliatura oral que é associada com a região da boca. Os cílios do corpo são iguais, mas são mais longos na extremidade posterior, daí o nome caudatum. Os cílios são organelas de locomoção e coleta de alimentos, eles também atuam como receptores sensoriais e detectam os estímulos do ambiente externo.

Ultra Estrutura de Cilia:
Os cílios e flagelos têm uma composição fibrilar. Na base o cilo tem o diâmetro de cerca de 0,2 mícron ou 2.000 A0 que podem estar até 10 mícrons acima da superfície celular. Os cílios são delimitados por uma membrana unitária de 90 A0 de espessura que se assemelha e permanece contínua com a membrana plasmática. O espaço delimitado do cílio contém uma substância aquosa conhecida como matriz.
Na matriz, permanecem onze fibrilas longitudinais ou microtubulosos embutidos. Das onze fibrilas, duas estão localizadas no centro, enquanto as nove fibrilas restantes permanecem dispostas de forma periférica em torno das fibrilas centrais. Cada uma das nove fibrilas externas tem um diâmetro de 360A0 e é composta por duas subfibrilas de 180 a 250A0 de diâmetro.
Estas sub-fibrilas são designadas como sub-fibril A e sub-fibril B.
ADVERTISEMENTOS:
A sub-fibril A é ligeiramente maior que a sub-fibril B. A sub-fibril A dá duas projecções espessas ou braços de um dos seus lados. Os braços da sub-fibrilha A de todas as fibras exteriores permanecem dirigidos no sentido horário. Além disso, a sub-fibril A ocorre mais perto do centro do cório do que a sub-fibril B. Ambas as sub-fibrilas têm uma parede comum de 50A0 de espessura.
As duas fibrilas centrais não têm sub-fibrilas pareadas como as nove fibrilas periféricas, mas cada uma contém apenas um único túbulo. Cada fibrilha central tem um diâmetro de cerca de 250A0 e é composta por uma parede de 60A0 de espessura.
As fibrilas centrais permanecem separadas por um espaço de 350A0 e permanecem envoltas numa bainha comum. Gibbnos (1967) relatou que a bainha das fibrilas centrais dá nove ligações ou raios orientados radialmente para cada sub-fibril A.

A microscopia eletrônica de alta resolução revelou que cada uma das fibrilas periféricas e centrais dos cílios e flagelos é composta de dez a doze filamentos de 40A0 de espessura. Cada filamento é feito com contas. Cada fio permanece disposto nas grades de 40 por 50A0 no plano da parede do túbulo. Estes talões são considerados como a subunidade básica da estrutura do tímpulo.
(iv) Sistema Infra-Cilíndrico:
O sistema infra-ciliar está localizado logo abaixo dos alvéolos peliculares. É constituído pelo cinetossoma ou corpo basal e cinetodésma. Os cílios surgem dos cinetosomas e de cada cinetosoma surge um delicado fibrilação citoplasmática chamado cinetodesma, (Fig. 20.2).
Deitado abaixo da película ligeiramente para a direita, mas unido a todos os cinetosomas de uma linha longitudinal, é um feixe longitudinal de vários cinetosomas, um cinetosoma de cada cinetosoma estende-se por uma distância anterior no seu próprio feixe de cinetosomas.
Uma linha longitudinal de cinetossomas com seus cinetossomas forma uma unidade longitudinal chamada de cinetossoma. Todos os cinetos ou cinécias formam o sistema infra-ciliar de um ciliado. Os cinécios encontram-se no córtex abaixo da película, o seu número é bastante constante para cada ciliado.
O sistema infra-ciliar controla e coordena os movimentos dos cílios, e provoca a formação de organelas na divisão celular, por exemplo, algumas cinécias formam a boca. Na fissão binária das ciliadas as cinécias são cortadas transversalmente em duas, cada uma indo para uma célula filha, isto é chamado de fissão perikinetal.
(v) Sulco Oral e Cytopyge:
No lado ventrolateral há uma grande depressão oblíqua e rasa chamada ranhura oral ou peristoma que dá ao animal uma aparência assimétrica. Corre obliquamente de um lado para trás (geralmente da esquerda para a direita, mas em alguns casos da direita para a esquerda) e termina um pouco atrás do meio do corpo. A ranhura oral leva a uma pequena depressão cónica em forma de funil chamada vestíbulo.
ADVERTISEMENTOS:
O vestíbulo leva directamente à abertura fixa em forma oval chamada cítromo (boca). Estendendo-se directamente do citótomo em direcção ao centro do corpo está a ampla citofaringe. A citofaringe vira-se então acentuadamente para o lado posterior para se tornar no esófago estreito e afunilado.
Assim, o esófago é aproximadamente paralelo à superfície do corpo do Paramecium excepto na sua extremidade posterior. Aqui o esôfago vira-se novamente para o centro do animal para conduzir ao vacúolo do alimento em formação.
A citopia (também chamada de ânus celular ou mancha anal ou citopropia) encontra-se na superfície ventral do corpo quase verticalmente atrás do citótomo ou boca. As partículas de alimentos não digeridas são eliminadas através da citopia. A citofaringe da citofaringe é muito complicada.
Gelei (1934) relatou a presença de quatro filas e Lund (1941) observou pelo menos quatro filas. Uma estrutura chamada penniculus é encontrada na parede esquerda da citofaringe e espiras através de aproximadamente 90 graus, de modo que sua extremidade posterior está na superfície oral (ventral) do esôfago.
De acordo com Lund, o penniculus consiste em oito filas de cílios dispostas em dois blocos bem definidos de cada. Uma faixa similar composta de quatro filas de cílios longos que são menos compactos do que no pennículo é chamada de quadrículo. Ela desce em espiral pela parede dorsal da cavidade bucal e termina perto do penículo.
O pennículo e o quadrículo foram erradamente chamados de membrana ondulada por alguns trabalhadores. O quadriculo e o pennículo controlam a passagem dos alimentos. Não se sabe como funcionam os cílios, provavelmente as suas fibrilhas contraem-se de forma rítmica o que provoca a dobra. Gelei (1925) apontou que a função do penniculus é a forçagem dos elementos alimentares para dentro do corpo.

(vi) Citoplasma:
O citoplasma é diferenciado em uma zona estreita, externa ou cortical chamada ectoplasma e uma região maior, interna ou medular chamada endoplasma.
(vii) Ectoplasma:
O ectoplasma (ectosarc ou córtex) é uma parte permanente do corpo, marcadamente delimitada do endoplasma. O ectoplasma forma uma camada externa firme, clara, fina e densa. Ele contém os tricocistos, cílios e estruturas fibrilares e é limitado externamente por uma cobertura chamada pelicular.
(viii) Tricocistos:
Embutidos no ectoplasma em ângulo recto com a superfície estão pequenos sacos em forma de fuso chamados tricocistos. Uma pequena mancha em cada margem anterior e posterior do hexágono marca a posição de um tricócito. Eles são preenchidos com um fluido refrativo, denso e com uma substância que causa inchaço, na extremidade externa é uma cabeça cônica ou espiga.
Provenientes de cinetosomas de cílios, migram e se localizam a igual distância no endoplasma. Quando o animal está irritado, os tricocistos são descarregados como longos fios pegajosos. Um tricócito descarregado tem um espigão opaco como uma unha invertida, e uma haste longa estriada, mas a haste não é vista no estado não descarregada e provavelmente se forma durante a descarga.
A função dos tricocistos é incerta, mas eles são descarregados como reação a contatos locais e lesões, eles podem servir como organelas de defesa.
But isto é incerto porque os tricocistos são ineficazes contra Didinium, o principal predador do Paramecium, eles podem ser para fixar o animal em um local durante a alimentação. Em alguns ciliados, os tricocistos agem como organelas de ofensa. Depois que os tricocistos são descarregados, regenerados a partir de cinetosomas.


(ix) Neuromotorium and Associated Fibrils:
De acordo com Lund (1933) na parede dorsal esquerda da citofaringe, aproximadamente ao nível da margem posterior do citótomo é uma massa muito pequena, bilobada, o neuromotorium. Do neuromotorium, as fibrilhas irradiam para o endoplasma.
Destas quatro ou mais geralmente passam quase para a parede dorsal do corpo, mas as demais são mais curtas e não estão definitivamente em posição. Todos são denominados fibrilas endoplasmáticas. A sua função é desconhecida, mas podem coordenar os movimentos de alimentação dos cílios orais. As fibrilhas também podem dar suporte mecânico, elasticidade, contratilidade, condutividade e influência metabólica.

(x) Endoplasm:
O endoplasma ou medula é a parte mais fluida e volumosa do citoplasma que contém muitos grânulos citoplasmáticos assim como outras inclusões e estruturas de natureza especializada. As inclusões citoplasmáticas são mitocôndrias, aparelhos de Golgi, vacúolos, cristais, grânulos e cromidias, etc. Outras estruturas, ou seja, núcleos, vacúolos contráteis e vacúolos alimentares também são encontrados no endoplasma.
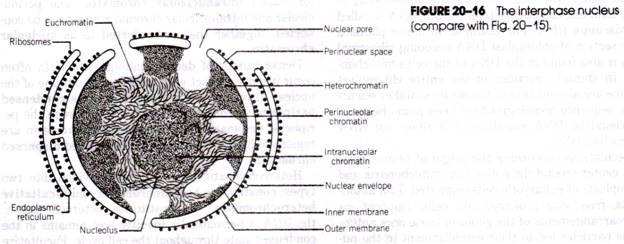
Núcleo:
No endoplasma perto do citoplasma estão dois núcleos, ou seja, o Paramecium é heterocariótico, um grande macronúcleo elipsoidal e granular e outros pequenos micronúcleos compactos.
Macronucleus é um corpo conspícuo, elipsoidal ou em forma de rim. É de um tipo compacto contendo fios finos e grânulos de cromatina discretos de tamanho variável e firmemente embalados e embutidos em uma matriz acromática. Possui muitos núcleos e muito mais material cromatinado (ADN).
É um núcleo somático ou vegetativo. Divide-se amitoticamente e controla as funções vegetativas (atividades metabólicas) do animal. Ele sofre de mitose.
Micronucleus é pequeno, compacto e esférico. É geralmente encontrado perto do macronúcleo, muitas vezes em uma concavidade. Os grânulos e fios de cromatina fina estão uniformemente distribuídos por toda a estrutura. O micronúcleo divide mitoticamente e controla a reprodução.
O número de micronúcleos varia com a espécie; é um em P. caudatum, dois em P. aurelia e muitos em P. multimicronucleatum. O micronúcleo contém um nucléolo distinto em P. aurelia mas não é encontrado em P. caudatum. Moisés (1949; 1950) relatou que o macronúcleo e o micronúcleo são idênticos em composição química.
(xi) Vacúolos contráteis:
Existem dois grandes vacúolos contráteis cheios de líquido, cada um situado perto de uma extremidade do corpo perto da superfície dorsal. A sua posição é fixa (ao contrário da Amoeba), situam-se entre o ectoplasma e o endoplasma, mas são organelas temporárias desaparecendo periodicamente. Em algumas espécies, parecem ter uma membrana de revestimento, caso em que não desaparecem completamente durante a sístole.
Conectados a cada vacúolo contrátil estão cinco a doze canais tubulares radiantes, cada um deles constituído por uma parte terminal, uma ampola longa que colapsa quando vazia, e um canal injetor curto que se abre para o vacúolo.

Os canais comunicam com uma grande parte do corpo de onde retiram os líquidos e os despejam no vacúolo que é, assim, reconstituído e cresce em tamanho, quando o vacúolo contrátil atinge o seu tamanho máximo contrai-se subitamente (sístole) e descarrega o seu conteúdo através de um poro permanente na película, depois os canais voltam a formar os vacúolos contráteis, os canais não desaparecem por completo, uma vez que são estruturas permanentes.
Os dois vacúolos contráteis descarregam irregularmente, o posterior contrai-se mais rapidamente porque está perto da citofaringe e mais água entra nela. A função principal dos canais e dos vacúolos contráteis é hidrostática, eles removem o excesso de água do protoplasma, a água é parcialmente absorvida e parcialmente absorvida durante a alimentação.
A substância do resíduo nitrogenado tem compostos de amônia e alguns uratos que são expelidos dos vacúolos contráteis junto com o CO2, mas não há evidência de que a matéria excretora seja secretada pelo protoplasma para os canais ou vacúolos contráteis.
Existe uma citoplasma fixa permanente ou anus abaixo do peristomo para um lado da citofaringe, através da qual são expelidos restos não digeridos de comida e uratos.
O estudo microscópico electrónico de vacúolos contráteis revelou que cada aparelho contrátil consiste em alguns dos túbulos de retículo endoplasmático, túbulos nefrídios, canais de alimentação, vacúolos acessórios (canais radiais) e vacúolos contráteis principais. Os aspiradores acessórios são supostamente as ampolas dos canais de alimentação (Fig. 20.11).
(xii) Aspúolos para alimentos:
Estes são corpos aproximadamente esféricos, não contráteis variando em tamanho e número no endoplasma. Eles contêm partículas de alimentos ingeridas, principalmente bactérias e uma pequena quantidade de fluido limitada por uma fina membrana definida. Volkonsky (1934) propôs o nome de gastriolas para estes vacúolos. Associados aos vacúolos alimentares estão os grânulos digestivos.
Locomotion of Paramecium Caudatum:
Paramecium Caudatum realiza a locomoção por dois métodos, a saber, metabolismo ou contorções corporais e por cílios.
(i) Metabolia ou Contorções Corporais:
O corpo do Paramecium Caudatum possui elasticidade, ele pode se espremer através de uma passagem mais estreita que seu corpo, após a qual o corpo assume sua forma normal. Esta alteração temporária da forma do corpo é metabólica, ela é trazida em Paramecium pelo protoplasma.
(ii) Locomoção Ciliar:
Locomotion trazida por cílios é o método principal. Os cílios podem bater para frente ou para trás permitindo que o animal nade anterior ou posteriormente.
Normalmente o animal nada para a frente, os cílios batem para trás mas obliquamente, os cílios endurecem e dobram para trás rapidamente para quase tocar a superfície do corpo, isto é chamado de AVC eficaz; então os cílios tornam-se coxos e voltam lentamente para a posição vertical original, isto é, chamado de AVC de recuperação.
Cílios da mesma linha transversal batem juntos e os da mesma linha longitudinal batem um após o outro da extremidade anterior para a posterior.

Este movimento coordenado de cílios é chamado de ritmo metacronal, que é devido ao sistema infra-ciliar; isto causa nadar para frente pelo animal. Mas quando os cílios do corpo estão batendo obliquamente para trás, então ao mesmo tempo os cílios mais longos da ranhura oral batem mais vigorosamente, o que faz com que a extremidade anterior gire para a esquerda.
A acção dos cílios do corpo e da ranhura oral faz o animal rodar no seu longo eixo. Esta rotação é sempre para a esquerda (excepto em P. calkinsi que gira em espiral à direita).
Esta combinação de movimento para a frente, oscilação e rotação faz com que o animal se desloque para a frente numa espiral no sentido contrário ao dos ponteiros do relógio. Esta trajetória tem um eixo reto, e a mesma superfície corporal do animal permanece em direção ao eixo da trajetória em espiral. Mas ao nadar para trás, todas as espécies giram para a direita.
A batida ciliar pode ser invertida para que os cílios se movam obliquamente para a frente pelo qual o animal nada para trás. Pela acção ciliar, o Paramecium move-se com uma velocidade de 1500 microns ou até mais por segundo.
Jennings alegou que a espiral de Paramecium se deve ao fato de que enquanto os cílios atacam principalmente para trás, eles o fazem obliquamente para a direita, fazendo com que o animal role para a esquerda.
Aquele desvio do corpo para a superfície aboral deve-se em grande parte ao maior poder do golpe efetivo dos cílios orais que atingem mais diretamente para trás. O resultado – a rotação do Paramecium no seu longo eixo – permite que o Paramecium siga um curso mais ou menos recto na formação de grandes espirais.
Nutrição do Paramecium Caudatum:
No Paramecium Caudatum, a nutrição é holozóica. Os alimentos são constituídos principalmente por bactérias e Protozoários Minutos. O paramécio não espera pelo alimento, mas caça activamente por ele.
É afirmado que o Paramecium Caudatum mostra uma escolha na selecção do seu alimento, mas parece não haver base para isso, embora envolva apenas certos tipos de bactérias; os dados disponíveis sugerem que 2 a 5 milhões de indivíduos de Bacillus coli são devorados por um único Paramecium em 24 horas. Também se alimenta de plantas unicelulares como algas, diatomáceas, etc., e pequenos pedaços de animais e vegetais.
Mecanismo de alimentação:
Quando o Paramecium Caudatum entra numa região de alimento abundante, vem para descansar. Alimenta-se apenas em repouso ou quando nada muito lentamente, nunca se alimenta quando nada rápido. O batimento de cílios da ranhura oral faz com que um vórtice cónico de água carregada de comida seja varrido para dentro da ranhura oral a uma distância anterior à extremidade anterior (Fig. 20.16).
As partículas de comida vão então para o vestíbulo de onde algumas partículas de comida são rejeitadas e lançadas para fora, mas outras passam para o citótomo.
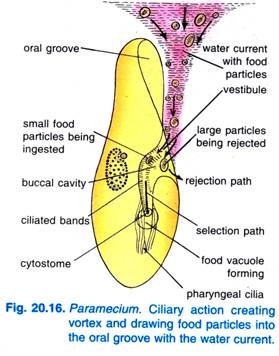
No final da citofaringe, forma-se um vacúolo de alimentos que se enche de partículas de alimentos. O quadrulus e o peniculi controlam a passagem dos alimentos para o vacúolo alimentar que se forma lateralmente. Quando o vacúolo alimentar atinge um determinado tamanho, as fibras pós-bucais prendem o vacúolo alimentar e este é apertado por elas e iniciado no seu curso.
O vacúolo contém alguma água além do alimento. Os movimentos rotativos de fluxo de endoplasma chamados ciclosclosis transportam os vacúolos de alimentos ao longo de um curso definido que é funcionalmente equivalente a um tracto digestivo.
O tracto começa do fim da citofaringe, depois para o lado posterior, depois para a frente para circular com o endoplasma, depois para a superfície dorsal, depois para a extremidade anterior, depois para baixo até à citoplasma. No início da sua viagem o vacúolo alimentar diminui de tamanho, depois aumenta novamente.

Digestão e ingestão de Paramecium Caudatum:
Durante a ciclosclosis, a digestão ocorre por enzimas secretadas por protoplasma nos vacúolos. Na digestão, as proteínas são transformadas em aminoácidos, os carboidratos em açúcares solúveis e glicogênio, e as gorduras são provavelmente também digeridas.
Os conteúdos dos vacúolos alimentares são inicialmente ácidos (pH cerca de 4) e depois tornam-se alcalinos, a maior digestão ocorre durante a fase alcalina. A matéria não digerida é ingerida através da citopatia com alguma força.
Cyclosis pode ser demonstrado experimentalmente; se o leite corado com vermelho Congo é alimentado com Paramecium, os glóbulos gordos do leite nos vacúolos alimentares tornar-se-ão primeiro vermelhos devido à reacção ácida das enzimas, depois mudarão de tons de roxo para azul devido à reacção alcalina, os vacúolos mostrarão o curso da ciclosclosis.
Respiração e Excreção de Paramecium Caudatum:
A troca de gases (oxigénio e dióxido de carbono) ocorre através da película semi-permeável como outros protozoários de água doce pelo processo de difusão. O Paramecium Caudatum obtém o seu oxigénio a partir da água circundante. O dióxido de carbono e resíduos orgânicos como a amônia resultante do metabolismo são provavelmente excretados pela difusão para fora na água no sentido inverso.

6. Osmoregulação no Paramecium Caudatum:
Paramecium Caudatum tem dois vacúolos contráteis, um anterior e um posterior. A função dos vacúolos contráteis é a osmoregulação, ou seja, regular o conteúdo de água do corpo e pode servir também na excreção de resíduos nitrogenados como uréia e amônia.
Excesso de água (devido à endosmose contínua) dentro do citoplasma é segregado nos túbulos do retículo endoplasmático e vai para os túbulos nefrídios → canais de alimentação → e recolhe em ampola de uma série de 6 a 11 canais radiantes que convergem para e descarregam em cada vacúolo. Os canais são mais evidentes quando um vacúolo está se formando.
Quando cada vacúolo é inchado (diástole) até um certo tamanho, contrai-se (sístole) e descarrega para o exterior provavelmente através de um poro. Os vacúolos contráteis contraem-se alternadamente, em intervalos de 10 – 20 segundos.
O vacúolo contrátil posterior funciona mais rapidamente que o anterior devido à entrada de grande quantidade de água na região posterior pela citofaringe. Os vacúolos contráteis mantêm uma concentração ótima de água no citoplasma do corpo através da eliminação do excesso.

Comportamento do Paramecium Caudatum:
As respostas do Paramecium Caudatum a vários tipos de estímulos são aprendidas através do estudo das suas reacções e do agrupamento ou dispersão dos indivíduos numa cultura. A resposta é positiva se o animal se move em direção a um estímulo e negativa quando ele se afasta. A um estímulo adverso, o animal continua a dar a reação evitadora até escapar.
Ao evitar a reação, a batida ciliar inverte-se, o animal retrocede uma curta distância, e depois gira em um caminho cônico, oscilando a extremidade anterior abortada enquanto gira na extremidade posterior. Todos os ajustes são feitos por tentativa e erro. As experiências mostraram que a extremidade anterior do animal é mais sensível do que as outras partes.
As respostas do Paramecium a diferentes estímulos podem ser agrupadas como se segue:
(i) Reacções ao contacto (Thigmotaxis):
As reações ao contato são variadas em Paramecium. Se a extremidade anterior for ligeiramente tocada com um ponto fino, ocorre uma forte reação evitadora. Quando um Paramecium nado colide com algum objeto na água, mas se tocado em outro lugar, pode não haver resposta. Um indivíduo em movimento lento frequentemente responde, positivamente ao contacto com um objecto, vindo a descansar sobre ele.
(ii) Reações a produtos químicos (Quimiotaxis):
Generalmente a Paramecia responde a um estímulo químico, evitando a reacção. Se uma gota de solução salina fraca (0,5 por cento) é introduzida numa população de Paramecium em um micro deslizamento, os animais respondem com a reação de evitar e nenhum entra na gota. Aos ácidos, no entanto, a resposta é positiva mesmo quando a concentração é de força suficiente para matá-los.
(iii) Reações à temperatura (Termotaxis):
Paramecium procura uma temperatura óptima de 24 a 28°C. Quando uma mudança de temperatura ocorre acentuadamente acima ou abaixo da faixa ideal, Paramecia mostra uma reação evitadora. Um maior calor estimula o movimento rápido e evita reacções até que os animais escapem ou sejam mortos.
(iv) Reacções à luz (Fototaxis):
Com a excepção da bursaria verde Paramecium bursaria, que é positivamente fototáctica, outras espécies são indiferentes à luz comum. No entanto, quando a intensidade da luz é repentina e acentuadamente aumentada, geralmente segue-se uma reacção negativa. A Paramecia apresenta uma resposta negativa imediata aos raios ultravioletas.
(v) Reações à corrente elétrica (Galvanotaxis):
Paramecia responde a estímulos eléctricos. Quando dois eletrodos são colocados um em frente ao outro em um prato raso contendo Paramecia e uma corrente constante aplicada, todos os organismos nadam na mesma direção em direção ao cátodo ou eletrodo negativo onde se concentram em grande número.
Se a direção da corrente elétrica for invertida enquanto o Paramecia nada em direção ao cátodo, os organismos invertem a direção e nadam em direção ao novo cátodo.

(vi) Reações à corrente de água (Reotaxis):
Paramecia mostra uma reotaxis positiva. Numa corrente de água suave o Paramecia irá mover-se principalmente com o fluxo com as suas extremidades anteriores a montante.
(vii) Reacções à gravidade (Geotaxis):
Paramecia geralmente exibe uma resposta negativa à gravidade como visto numa cultura onde muitos indivíduos se juntam sob a película de superfície com as suas extremidades anteriores mais acima. Se Paramecia são introduzidos num tubo em U com enchimento de água invertida, com as duas extremidades em forma de U, eles movem-se imediatamente para cima, para a parte horizontal do tubo.
Reprodução em Paramecium Caudatam:
O Paramecium Caudatum reproduz-se assexualmente por fissão binária transversal e também sofre vários tipos de reorganização nuclear, tais como conjugação, endomixis, autogamia, citogamia e hemixis, etc.
(i) Fissão Binária Transversa:
Fissão binária transversa é o tipo mais comum de reprodução assexuada em Paramecium. É um processo assexual claramente único, no qual um espécime totalmente cultivado se divide em dois indivíduos filhos sem deixar um cadáver parental.
O plano de divisão é através do centro da célula e em um plano em ângulo reto com o longo eixo do corpo. A divisão do corpo celular como um todo é sempre precedida pela divisão dos núcleos; de fato, parece que a reprodução é iniciada pela atividade nuclear e pela divisão.

Paramecium Caudatum reproduz-se por fissão binária transversal em condições favoráveis. Na fissão binária, o micronúcleo se divide por mitose em dois micronúcleos filhos, que se movem para extremos opostos da célula. O macronúcleo se alonga e se divide transversalmente por amitose.
Outra citofaringe é desprendida e aparecem dois novos vacúolos contráteis, um perto da extremidade anterior e outro perto da extremidade posterior. Entretanto, um sulco de constrição aparece perto do meio do corpo e se aprofunda até que o citoplasma esteja completamente dividido.
As duas paramecias “filhas” resultantes são de tamanho igual, cada uma contendo um conjunto de organelas celulares. Das duas paramecias filhas produzidas, a anterior é chamada de proter e a posterior é chamada de opisthe. Elas crescem em tamanho real antes que outra divisão ocorra.
O processo de fissão binária requer cerca de duas horas para ser concluído e pode ocorrer uma a quatro vezes por dia, produzindo 2 a 16 indivíduos. Cerca de 600 gerações são produzidas em um ano.
A taxa de multiplicação depende das condições externas da alimentação, temperatura, idade da cultura e densidade populacional; também dos fatores internos de hereditariedade e fisiologia. Naturalmente, se todos os descendentes de um indivíduo sobrevivessem e se reproduzissem, o número de paramecias produzidas logo seria igual ao volume da terra.
O termo clone é usado para se referir a todos os indivíduos que foram produzidos a partir de um indivíduo por fissão. Todos os membros de um clone são hereditários.
(ii) Conjugação:
Ordinariamente o Paramecium Caudatum se multiplica por fissão binária por longos períodos de tempo, mas em intervalos de tempo isto pode ser interrompido pela união de dois animais ao longo de suas superfícies orais para o processo sexual de conjugação.
A conjugação é definida como a união temporária de dois indivíduos que trocam mutuamente micro material nuclear. É um tipo único de processo sexual no qual dois organismos se separam logo após a troca de material nuclear.
Sonneborn (1947), com base no comportamento de acasalamento do Paramecium Caudatum, tem relatado que cada espécie de Paramecium existe em várias variedades ou syngens. Além disso, dentro de cada syngen há um número de tipos de acasalamento geralmente dois.
Os tipos de acasalamento permanecem morfologicamente idênticos, mas apresentam diferenças fisiológicas. Em P. aurelia, há 14 syngens e 28 tipos de acasalamento, enquanto em P. caudatum, há 16 syngens e 32 tipos de acasalamento. Foram feitas observações que normalmente não se conjugam com membros do seu próprio tipo de acasalamento nem com as outras variedades, mas apenas com o segundo tipo de acasalamento da sua própria variedade.

Factores Indutores da Conjugação:
Os factores que induzem a conjugação variam de espécie para espécie, mas alguns deles são dados abaixo:
1. A conjugação ocorre geralmente sob condições de vida desfavoráveis; diz-se que a fome ou escassez de alimentos e dieta bacteriana particular ou certos produtos químicos induzem o processo de conjugação em certas espécies de Paramecium.
2 A conjugação ocorre após cerca de 300 gerações assexuadas de fissão binária, ou alterna com a fissão binária em longos intervalos para rejuvenescer o clone moribundo, ou seja, ocorre nos indivíduos que devem ter passado por um número desejável de gerações assexuadas, dito ser o período de imaturidade, e depois tornam-se sexualmente maduros para se conjugarem.
4. Diz-se que a escuridão repentina em condições de luz e baixas temperaturas induz o processo de conjugação em algumas espécies.
5. A conjugação não ocorre durante a noite ou escuridão; ela começa de manhã cedo e continua até a tarde.
6. Diz-se que uma substância proteica nos cílios de indivíduos do tipo acasalamento induz a conjugação.
Processo de Conjugação:
O processo de conjugação difere em diferentes espécies de Paramecium, mas o relato subnotificado diz respeito ao processo de conjugação de P. caudatum (Fig. 20.21).
Em conjugação dois Paramecium caudatum (referidos como pré-conjugantes) dos tipos de conjugação opostos da mesma variedade se juntam com suas superfícies ventrais e se unem por seus sulcos orais; seus cílios produzem uma substância na superfície do corpo que causa a adesão das duas paramecias conjugadas.
Deixam de se alimentar e o seu aparelho de ranhuras orais desaparece. A película e o ectoplasma, no ponto de contacto, de ambos quebram, e forma-se uma ponte protoplasmática entre os dois animais. Agora, estes indivíduos são chamados de conjugais.
Nesta condição, o par conjugador nada ativamente e simultaneamente uma série de mudanças nucleares acontecem em cada conjugador como descrito abaixo:
O macronúcleo começa a desintegrar-se, torna-se solto em textura e forma uma meada torcida complexa, durante a última metade do período de conjugação desaparece finalmente sendo absorvido no citoplasma. O micronúcleo de cada conjugado se divide duas vezes, sendo uma delas uma divisão redutora.
Assim, quatro micronúcleos haplóides filhas são produzidos em cada conjugal. Três destes quatro micronúcleos degeneram em cada um, de modo que apenas um permanece.
Os restantes micronúcleos de cada conjugal dividem-se mitoticamente em dois pronúcleos desiguais ou núcleos cinegéticos formando um pro-núcleo feminino estacionário maior e um pro-núcleo masculino migratório menor e activo.
O pró-núcleo migratório de um conjugado atravessa a ponte protoplasmática e funde-se com o pró-núcleo estacionário de outro conjugado para formar um núcleo sinkaryon ou conjugação em que o número diplóide de cromossomas é restaurado e tem havido uma troca de material hereditário.
O processo foi comparado com a fertilização em animais superiores, mas isto não é fertilização porque não estão envolvidos gametas. Os conjugantes agora (após cerca de 12-48 horas) separam-se e são chamados ex-conjugantes. O sinkaryon de cada ex-conjugante se divide três vezes para formar oito micronúcleos em cada ex-conjugante.
Quatro dos oito micronúcleos aumentam e se tornam macronúcleos, e três dos outros quatro micronúcleos desaparecem.
Os restantes micronúcleos dividem-se e ao mesmo tempo o ex-conjugante divide-se por fissão binária em duas células, cada uma com dois macronúcleos e um micronúcleo. As células e seus micronúcleos dividem-se uma segunda vez para formar quatro paramecias de cada ex-conjugante, de modo que cada um tenha um macronúcleo e um micronúcleo.
Os novos macronúcleos, como também os micronúcleos, foram feitos de novo material. Estes novos núcleos provavelmente contêm um potencial novo e diferente que se reflecte nos indivíduos saudáveis.
Significado de Conjugação:
Um clone desaparecerá se a reorganização nuclear não ocorrer, mas o clone pode ser rejuvenescido para recuperar o seu antigo vigor através da reorganização nuclear, esta reorganização nuclear é trazida pela conjugação, assim, a conjugação é essencial para a fissão binária contínua.
O significado da conjugação foi resumido abaixo:
1. A conjugação serve como um processo de rejuvenescimento e re-organização através do qual a vitalidade da raça é restaurada. Se a conjugação não ocorre por longos períodos, a paramecia enfraquece e morre. (Woodruffs alega manter a paramecia saudável por 22.000 gerações sem conjugação é desmentida por Sonneborn, porque ele mostrou que toda a paramecia Woodruffs pertencia ao mesmo tipo de acasalamento).
2. Não há distinção de sexo nos conjugados, embora apenas a paramecia de dois tipos diferentes de acasalamento da mesma variedade irá conjugar.
3. Não há distinção de sexo, no entanto o pro-núcleo migratório ativo é considerado como masculino e o pro-núcleo estacionário como o feminino.
4. A conjugação é apenas uma união temporária, não há fusão de citoplasma e não se produz zigoto, mas o núcleo de cada ex-conjugante contém material hereditário de dois indivíduos conjugados.
5. A conjugação traz a substituição do macronúcleo por material do sinkaryon, este é um evento de fundamental importância. Na fissão binária os cromossomos do macronúcleo foram distribuídos aleatoriamente para as células filhas, a fissão binária contínua tornou o clone fraco com algumas anormalidades estruturais.
A conjugação traz a formação do número correto de cromossomos no macronúcleo, para que a raça seja renovada em vigor. O papel do micronúcleo é restaurar um cromossoma equilibrado e um complexo genético.
Comportamento Aberrante na Reprodução em Paramecium Caudatum:
Paramecium Caudatum mostra certas variações no seu comportamento nuclear durante a fissão e conjugação, estes desvios são endomixis, autogamia, citogamia e hemixis. Nos três primeiros processos é feita a recombinação genética e um novo macronúcleo é formado a partir do micronúcleo.
(i) Endomixis:
Woodruff e Erdmann (1914) primeiro de tudo relataram um novo processo de reorganização nuclear, endomixis (Gr., endon = dentro; mixis = mistura) em Paramecium aurelia, uma espécie bimicronucleada (Fig. 20.22). Este processo foi descrito como ocorrendo periodicamente no qual um novo aparelho macro-nuclear é produzido sem formação de sinkaryon. A endomixia ocorre dentro de um único indivíduo.
De acordo com Woodruff e Erdmann, o macronúcleo degenera e o micronúcleo se divide duas vezes para formar oito micronúcleos. Seis micronúcleos se degeneram e dois permanecem. Com apenas dois micronúcleos restantes, o animal se divide por fissão em duas células, cada uma com um micronúcleo.
Os micronúcleos de cada célula filha dividem-se duas vezes para formar quatro micronúcleos. Dois micronúcleos aumentam para formar macronúcleos. O animal e seus micronúcleos se dividem para formar dois indivíduos filhos, cada um tendo um macronúcleo e dois micronúcleos.
Endomixis ocorre naquela variedade de P. aurelia que não conjuga, portanto, o efeito de endomixis pode ser o mesmo que o da conjugação, já que ambos os processos trazem a substituição do macronúcleo por material do micronúcleo, e ambos os processos rejuvenescem a vitalidade da raça.
Mas os dois processos diferem porque não há fusão de pronúcleos em endomixis; endomixis pode ser comparado a partenogênese.
No entanto, alguns trabalhadores têm afirmado com boas razões que o endomixis não é um processo válido e tem sido descrito devido à observação deficiente. Com toda a probabilidade a endomixis não ocorre e pode ser apenas um caso especializado de autogamia.

Later, Erdmamm e Woodruff (1916) reportaram endomixis em Paramecium caudatum. Diller, entretanto, não acredita na validade deste processo e sente que Erdmann e Woodruff simplesmente combinaram os estágios de hemixis e autogamia em um esquema, endomixis.
(ii) Autogamia:
Diller (1934, 1936) e Sonneborn (1950) descreveram um processo de auto-fertilização ou autogamia ocorrendo em um único indivíduo em Paramecium aurelia (Fig. 20.23). Ele relatou que na autogamia três divisões micro-nucleares (pré-gâmicas), envolvendo maturação, produzem os núcleos cinegéticos (pronúcleos).
Durante a autogamia em P. aurelia, os dois micronúcleos dividem-se duas vezes (uma vez a meio) para formar oito micronúcleos, seis dos quais degenerados. Enquanto isso o macronúcleo cresce em massa semelhante a meada que se parte em pedaços mais tarde para ser absorvido no citoplasma. Dois dos oito micronúcleos, como pronúcleos, entram em um cone protoplasmático saliente próximo à boca da célula.
Os dois pronúcleos fundem-se para formar o sinkaryon. O sinkaryon divide-se duas vezes para formar quatro micronúcleos. Dois micronúcleos tornam-se macronúcleos. O Paramecium e seus micronúcleos se dividem para formar dois indivíduos filhos, cada um com um macronúcleo e dois micronúcleos. Este processo é concluído em cerca de dois dias.
Autogamia traz o rejuvenescimento da raça.
>
Forma-se em conjugação na medida em que o novo macronúcleo é formado por material do micronúcleo, no novo macronúcleo o número correto de cromossomos é restaurado; e também no fato de que a fusão de dois pronúcleos ocorre. Mas a autogamia difere da conjugação porque apenas um indivíduo participa da autogamia e ela fornece ambos os pronúcleos, é uma espécie de auto fertilização.
(iii) Hemixis:
Diller (1936) relatou a hemixis em Paramecium aurelia (Fig. 20.24). A hemixis é principalmente um processo de fragmentação e divisão macro-nuclear sem qualquer atividade micro-nuclear incomum. Diller classificou a hemixis em quatro tipos: A, B, C e D como mostrado na Fig. 20.24 em P. aurelia, mas ele também encontrou todos os tipos em culturas em massa de P. caudatum e P. multimicronucleatum.
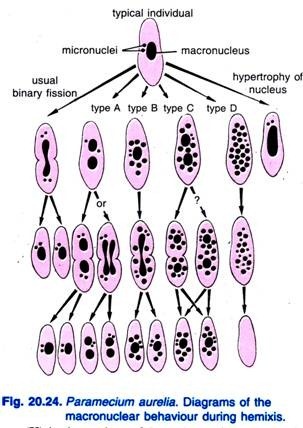
Type A é a forma mais simples de hemixis caracterizada por uma divisão do macronúcleo em duas ou mais partes. Esta divisão não está sincronizada com a divisão micro-nuclear.
Tipo B é caracterizado pela extrusão de uma a 20 ou mais bolas de cromatina do macronucleus para o citoplasma.
O tipo C caracteriza-se pela divisão simultânea do macronúcleo em duas ou mais porções principais e a extrusão de esferas macro-nucleares no citoplasma.
Tipo D é considerado como representando condições patológicas nas quais o macronúcleo sofre fragmentação completa em bolas de cromatina que eventualmente desaparecem da célula. Micronúcleos geralmente desaparecem antes da dissolução do macronúcleo.
(iv) Citogamia:
Wichterman (1939) relatou outro processo sexual no Paramecium caudatum, que ele chamou de citogamia. Na citogamia, não há troca nuclear. Neste processo, dois indivíduos se unem por suas superfícies ventrais, mas a película dos dois indivíduos não se rompe.
O micronúcleo de cada indivíduo divide-se três vezes para formar oito micronúcleos, seis dos quais se desintegram em cada indivíduo. Os dois micronúcleos restantes se fundem para formar um sinkaryon em cada célula. Os animais agora se separam.
Citogamia difere da autogamia por haver dois animais em contacto um com o outro, mas assemelha-se à autogamia e conjugação na fusão de dois pronúcleos. A citogamia difere da conjugação na medida em que não há troca nuclear entre os dois animais que se juntam.
Particulas Citoplasmáticas Relatadas em Paramecium Caudatum:
(i) Kappa Particles:
Em 1938, T.M. Sonneborn relatou que algumas raças (conhecidas como assassinas ou estirpe assassina) de Paramecium produzem uma substância venenosa, chamada paramecina, que é letal para outros indivíduos chamados sensitivos. A paramecina é solúvel em água, difusível e depende para a sua produção de algumas partículas localizadas no citoplasma do Paramecium (estirpe assassina).
Estas partículas são chamadas de partículas kappa. As partículas kappa têm DNA e RNA. Um paramécio killer pode conter centenas de partículas kappa. O estudo detalhado destas partículas revelou que um gene dominante (K) no núcleo do Paramecium é necessário para que as partículas kappa existam, se multipliquem e produzam paramecina.
(ii) mµ, Partículas:
R.W. Siegel (1952) relatou outro tipo de partículas assassinas no citoplasma de algum Paramecium. Um Paramecium com partículas mp é chamado mate killer porque quando se conjuga com um Paramecium sem quaisquer partículas mµ chamadas mate sensitive, então mata as últimas. As partículas de mp também são compostas de DNA, RNA, etc.
Estas partículas existem apenas naquelas paramecias cujos micronúcleos contêm pelo menos um gene dominante de um dos dois pares de genes cromossómicos não ligados (M1 e M2).
(iii) Partículas de Pi:
Estas partículas são supostamente a forma mutante das partículas kappa mas não produzem qualquer tipo de substância venenosa.
(iv) Partículas de Lambda:
Estas partículas são relatadas em paramecia assassina e dizem produzir alguma substância responsável por causar lise ou desintegração da paramecia sensível, ou seja, que não a possuem.