- Introducere
- Materiale și metode
- Subiecți și paradigmă experimentală
- Imagistica prin rezonanță magnetică funcțională și semnalele dependente de nivelul de oxigen din sânge
- Selectarea regiunilor de interes pentru semnalele dependente de nivelul de oxigen din sânge provenite de la cerebel / trunchiul cerebral
- Calcularea undelor medii dependente de nivelul de oxigen din sânge, de intervalul de bătaie la bătaie și de respirație
- Rezultate
- Discuție
- Coincidența undelor de respirație și artefactul respirator dependent de nivelul de sânge-oxigen
- Activare neuronală ritmică asociată cu oscilația neuronală dependentă de nivelul de oxigen din sânge
- Activitatea neuronală acționează ca „forță motrice” pentru oscilațiile lente ale ritmului cardiac
- Limitări și perspective viitoare
- Concluzie
- Disponibilitatea datelor
- Declarație etică
- Contribuții ale autorilor
- Declarație privind conflictul de interese
- Recunoștințe
- Abbreviații
Introducere
Aritmia sinusală respiratorie (RSA) reflectă accelerarea ritmului cardiac (FC) în timpul inspirației și decelerarea FC în timpul expirației. Este un principiu fundamental (Yasuma și Hayano, 2004) și fenomenul central al respirației de rezonanță ritmată la 6/min asociat cu amplificarea variabilității frecvenței cardiace joase (HRV), cu o bunăstare emoțională sporită (Mather și Thayer, 2018) și cu o mai bună procesare a emoțiilor negative (Zaccaro et al., 2018). În mod interesant, există excepții de la RSA cu creșteri ale intervalului HR beat-to-beat (RRI) în timpul inspirației și cu faza RRI care conduce în raport cu ritmul respirator (Rassler et al., 2018). Această RSA paradoxală neobișnuită a fost găsită la o minoritate de participanți sănătoși în timpul scanării imagisticii prin rezonanță magnetică funcțională (fMRI), o situație inconfortabilă, uneori claustrofobă, asociată de obicei cu o anxietate crescută a stării (Munn et al., 2015; Pfurtscheller et al., 2018).
Undele de respirație spontană lentă cu o rată între 6 și 9 respirații/min sunt, de asemenea, însoțite de o HRV ridicată și ar putea facilita astfel procesarea emoțiilor neplăcute, cum ar fi anxietatea și stresul (Thayer și Lane, 2009). Datorită rolului principal al RRI asupra oscilațiilor respiratorii în timpul încetării neobișnuite a RSA (Rassler et al., 2018), pare probabil un oscilator neuronal autonom (pacemaker central; Julien, 2006) în creier, care acționează ca o sursă pentru oscilațiile lente ale RRI. Este demn de remarcat faptul că Perlitz și colab. (2004) au raportat un nou tip de ritm cardiovascular în banda de 0,15-Hz la om și câine, cu o origine comună în trunchiul cerebral. Cea mai interesantă caracteristică a acestui „ritm de 0,15-Hz” în bandă largă (care include și componente de frecvență apropiate de 0,1 Hz) este că perioadele de unde fusiforme sunt cuplate în fază cu respirația într-un raport de 1:1. Principalul nostru interes de cercetare a fost să identificăm acest stimulator cardiac, despre care se sugerează că ar fi localizat în trunchiul cerebral (Lambertz și Langhorst, 1998; Perlitz și colab, 2004), prin analiza semnalelor BOLD.
Semnalul BOLD nu cuantifică în mod direct activitatea neuronală în sine, dar este sensibil la modificări ale ratei metabolice cerebrale, ale fluxului sanguin cerebral și ale volumului sanguin cerebral (Obrig et al., 2000; Buxton et al., 2004) și la mai multe tipuri de mișcare (mișcări ale pieptului și ale vaselor de sânge; Birn et al., 2006). Prin urmare, semnalele BOLD pot fi compuse din componente neuronale și non-neuronale (de exemplu, mișcarea vaselor). Este de așteptat ca două semnale BOLD de origine diferită să poată fi identificate în trunchiul cerebral cu vasul său de sânge mare (artera bazilară) și sursa neuronală postulată de 0,15-Hz (Perlitz et al., 2004). Presupunem că unul dintre ele rezultă din mișcarea vaselor de sânge (BOLDv) și indică începutul unei acțiuni respiratorii lente, deoarece apare aproape simultan cu activarea neuronală a neuronilor respiratori. Celălalt se presupune că este asociat cu activarea neuronală (BOLDn) întârziată de timpul de cuplare neurovasculară de 2-3 s (Mateo et al., 2017). Speculăm că ambele semnale BOLD indică un stimulator cardiac central în trunchiul cerebral.
Materiale și metode
Subiecți și paradigmă experimentală
Grupul de subiecți studiați a fost același (vârsta de 23,8 ± 3,3 ani) ca cel analizat recent de Rassler et al. (2018). Înregistrarea și preprocesarea ECG și a respirației au fost descrise în acea lucrare. Toți participanții și-au dat consimțământul scris în cunoștință de cauză pentru protocolul studiului, care a fost aprobat de Comitetul local de etică de la Universitatea din Graz.
Imagistica prin rezonanță magnetică funcțională și semnalele dependente de nivelul de oxigen din sânge
Imaginile funcționale au fost achiziționate cu un scaner 3 T (Magnetom Skyra) utilizând o secvență GE-EPI multibandă (Moeller et al, 2010) cu o achiziție simultană în șase benzi cu TE/TR = 34/871 ms, unghi de flip de 52°, dimensiune voxel de 2 mm × 2 mm × 2 mm, 66 de felii axiale contigue (11 × 6), matrice de achiziție de 90 × 104 și un FOV de 180 mm × 208 mm. Această rată de scanare de 871 ms (frecvență de eșantionare de 1,15 Hz) permite studierea oscilațiilor lente în intervalul 0,1-0,15 Hz. Pentru mai multe detalii despre preprocesare, a se vedea Pfurtscheller et al. (2018). Atlasul de etichetare anatomică automată (AAL) (Tzourio-Mazoyer et al., 2002) a fost utilizat pentru a extrage cursurile de timp ale semnalelor BOLD în 116 regiuni de interes (ROI). Acest atlas nu prezintă ROI-uri în trunchiul cerebral, ci doar în imediata apropiere a cerebelului și a vermisului. Datorită proximității acestor regiuni și datorită contiguității unei artere majore, este rezonabil să presupunem că semnalul colectat din ROI-urile AAL etichetate „cerebel” reflectă parțial activitatea trunchiului cerebral.
Selectarea regiunilor de interes pentru semnalele dependente de nivelul de oxigen din sânge provenite de la cerebel / trunchiul cerebral
Printre cele 116 ROI ale atlasului AAL, ROI 91 până la ROI 108 corespund unor felii axiale din cerebel, inclusiv trunchiul cerebral (Tzourio-Mazoyer et al., 2002). Imaginile T1 din felia axială (spațiul Talairach z = -34) documentează proximitatea cerebelului și a trunchiului cerebral (Figura 1). Din cauza numărului mic de voxeli (<200), ROI-urile 107 și 108 au fost excluse din analiza ulterioară. ROI-urile din cerebel / trunchiul cerebral au fost inspectate vizual pentru a căuta ROI-uri adecvate, adică ROI-uri cu un raport semnal/zgomot bun și unde BOLD lente dominante. Au fost identificate următoarele ROI-uri: 96, 98, 100, 103 și 105. Numerele indică etichetele ROI în conformitate cu atlasul AAL. Numerele pare și impare denotă emisferele stângă și, respectiv, dreaptă.
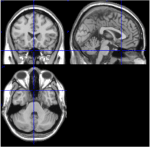
Figura 1. Imagini T1 din felii mediale, sagitale și axiale de la un subiect (spațiul Talairach 0, 20, -34). În felia sagitală (imaginea din dreapta sus), ponsul este clar vizibil, iar în felia axială (în partea de jos) este documentată proximitatea cerebelului și a trunchiului cerebral.
Calcularea undelor medii dependente de nivelul de oxigen din sânge, de intervalul de bătaie la bătaie și de respirație
Medierea permite îmbunătățirea raportului semnal-zgomot, dar necesită utilizarea unui declanșator. Deoarece nu există astfel de declanșatori în datele privind starea de repaus, au fost utilizate maximele (vârfurile) care apar ritmic ale semnalului RRI (Pfurtscheller et al., 2017): În primul rând, au fost identificate cele mai proeminente vârfuri ale semnalelor RRI, distanțate la cel puțin câteva secunde între ele. În cazul oscilațiilor de 0,1-Hz, aceste intervale între vârfuri au fost de aproximativ 10 s, iar în cazul oscilațiilor de 0,15-Hz, acestea au fost de aproximativ 7 s (un exemplu este ilustrat în figura 2 prin liniile verticale punctate aproape echidistante). Vârfurile RRI marcate au fost folosite ca declanșatori pentru calcularea mediei (epoci cu 6 s înainte și 6 s după declanșare) între semnalele BOLD, RRI și respirație.
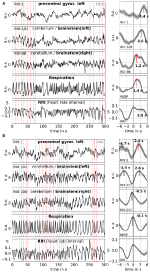
Figura 2. Exemple de semnale BOLD, RRI și de respirație în curs de desfășurare și undele medii corespunzătoare (±SE) pentru doi subiecți, unul cu oscilații dominante de 0,1-Hz și unul cu oscilații dominante de 0,15-Hz . Liniile verticale punctate din panourile din partea stângă indică maximele (vârfurile) oscilațiilor RRI utilizate ca declanșator pentru calcularea mediei. Vârfurile undelor mediate sunt indicate în panourile din partea dreaptă.
Rezultate
Oscilațiile BOLD lente în cerebelul/tezaurul cerebral se caracterizează prin calitatea favorabilă a semnalului lor, dar și mai important, prin coincidența lor cu oscilațiile în RRI și respirație. După cum s-a raportat în Rassler et al. (2018), oscilațiile de 0,1-Hz au predominat în timpul a ~37% din timpul de înregistrare, în timp ce oscilațiile de 0,15-Hz au predominat în ~45%. Două exemple caracteristice pentru oscilațiile de 0,1-Hz (subiectul 1Rb1) și 0,15-Hz (subiectul 11Ra1) sunt afișate în figura 2. În ambele cazuri, oscilațiile RRI de magnitudine mare coincid cu respirația sub forma unei creșteri a RRI în timpul inspirației și sunt, de asemenea, clar reflectate în semnalele BOLD din trunchiul cerebral. Rețineți, dinamica variabilă a oscilațiilor spontane și, respectiv, a undelor unice, în stare de repaus, oferă dovezi puternice că există nu doar unul, ci o varietate de ritmuri cardiovasculare cu componente de frecvență apropiate de 0,1 și 0,15 Hz. Unele dintre aceste ritmuri sunt amplificate în timpul procesării anxietății, altele nu, iar unele sunt sincronizate în cortexul prefrontal și în trunchiul cerebral, iar altele nu.
În figura 2 sunt prezentate exemple de semnale spontane și de unde medii de la doi subiecți caracteristici. Undele medii cu latențe de vârf marcate (distanța de la vârful RRI) ale tuturor subiecților sunt afișate în figura 3. De remarcat, variația relativ mică a undelor BOLD mediate în trunchiul cerebral subliniază fiabilitatea măsurătorilor de vârf.
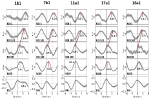
Figura 3. Undele medii (±SE) ale semnalelor BOLD din girusul precentral stâng (ROI 1), trunchiul cerebral stâng (BOLDn, ROI 103, 105), trunchiul cerebral drept (BOLDv, 96, 98, 100), respirația și intervalele RR (de sus în jos) de la toți cei cinci subiecți. Sunt marcate latențele de vârf (diferența față de vârful RRI) de la vârfurile importante. Decalajul temporal de 2-3 s între cele două semnale BOLD din cerebel/ trunchiul cerebral este indicat printr-o linie punctată.
Unda BOLD din emisfera dreaptă care precede unda de respirație cu 0,3 ± 0,2 s a fost interpretată ca BOLDv, iar unda BOLD din emisfera stângă care întârzie unda de respirație cu 2,3 ± 0,5 s a fost luată ca BOLDn. Diferența maximă între undele BOLDv și BOLDn a fost de 2,6 ± 0,4 s.
Undele medii din figura 3 evidențiază defazajul clar între cele două semnale BOLD din trunchiul cerebral (BOLDn, BOLDv) la toți cei cinci subiecți. Diferențele de vârf sunt indicate prin linii punctate. În plus față de cele două semnale BOLD din trunchiul cerebral, semnalul BOLD din girusul precentral stâng (ROI 1) a fost analizat ca și control. Deși undele BOLD medii din cortexul prefrontal (PFC) prezintă o varianță ridicată (SE), acestea se potrivesc în mod surprinzător de perfect cu undele BOLDn din cerebel / trunchi cerebral. Acest lucru confirmă interacțiunea puternică dintre PFC și trunchiul cerebral în gama de frecvențe lente în jurul valorii de 0,1 Hz.
Discuție
Coincidența undelor de respirație și artefactul respirator dependent de nivelul de sânge-oxigen
Respirația este însoțită nu numai de mișcarea toracelui, ci și de mișcarea vaselor de sânge cerebrale (Birn et al., 2006). A fost observată o decalare temporală stabilă de 0,3 ± 0,2 s între vârful pozitiv al semnalului BOLDv în cerebelul drept/tronc cerebral și maximul fiecărei unde de respirație (începutul expirației). Respirația a fost spontană, iar vârful respirației a întârziat față de vârful BOLDv, indicând, probabil, o vasomoție rapidă a arterei bazilare, un vas de sânge mare aproape de partea rostrală a ponsului. Această vasomoție ar putea fi considerată un artefact respirator indus de o modulare respiratorie a neuronilor simpatoexcitatori din nucleul reticular rostroventrolateral. O influență marcantă a generatorului central al modelului respirator asupra activității nervilor simpatici a fost observată la multe specii, inclusiv la om (Haselton și Guyenet, 1989; Häbler și Jänig, 1995; Eckberg, 2003; Mandel și Schreihofer, 2006). Scăderea activității acestor neuroni din măduva rostroventrolaterală (adică depresia simpatică) în timpul inspirației și o reintensificare bruscă a activității odată cu încetarea exploziilor frenice este unul dintre modelele predominante de cuplare respiratorie-simpatică (Haselton și Guyenet, 1989). Aceste oscilații simpatice modulate de respirație determină o vasodilatație maximă la sfârșitul inspirației (i.e, maximul semnalului BOLD imediat înainte de începerea expirației) și vasoconstricție în timpul expirației (minimul semnalului BOLD chiar înainte de debutul inspirației).
Activare neuronală ritmică asociată cu oscilația neuronală dependentă de nivelul de oxigen din sânge
Oscilațiile BOLD (BOLDn) au întârziat artefactul respirator periodic (BOLDv) cu 2.6 ± 0,4 s. Ambele oscilații BOLD au fost înregistrate simultan în felii axiale, BOLDn fiind asociat cu activitatea neuronală ca forță motrice a respirației spontane, iar BOLDv reflectând o mișcare vasculară asociată cu începutul expirației. Această întârziere de 2-3 s corespunde timpului de cuplare neurovasculară (Mateo et al., 2017).
Diversele influențe externe asupra emoției provocate de poziția incomodă în decubit dorsal cu capul subiectului într-un spațiu zgomotos limitat pot activa mai întâi zonele senzoriale primare și pot fi urmate de proiecția corticală către neuronii respiratori din trunchiul cerebral. Mai mult decât atât, influențele interne, comportamentale care decurg din centrele superioare modifică modelele metabolice de respirație. Ieșirea respiratorie finală implică o interacțiune complexă între structurile corticale, trunchiul cerebral și sistemul limbic și este caracteristică pentru respirația emoțională (Homma și Masaoka, 2008; Kato et al., 2018). Prin urmare, este destul de plauzibil ca oscilațiile din girusul precentral stâng (ROI 1) să fi fost sincronizate cu oscilațiile BOLD lente, induse neuronal în cerebel / trunchiul cerebral.
Activitatea neuronală acționează ca „forță motrice” pentru oscilațiile lente ale ritmului cardiac
Dinamica variabilă a modelelor de cuplare observate în starea de repaus indică o varietate de ritmuri cardiovasculare în banda de frecvență între 0,1 și 0,15 Hz. Acest lucru subliniază importanța, flexibilitatea și complexitatea interacțiunii dintre creier și inimă și merită cercetări intensive în continuare.
La toți cei cinci participanți, modelul de cuplare dintre respirație și HR a fost destul de contrar RSA tipic (Rassler et al., 2018), și anume RRI a crescut în timpul inspirației și a scăzut în timpul expirației. În timpul situațiilor provocatoare de anxietate, respirația tinde să fie mai rapidă și activitatea vagală diminuată, ceea ce duce la o HR mai mare (RRI mai scurt). Cu toate acestea, atunci când respirația este încetinită în mod inconștient/autonomic într-o situație provocatoare de anxietate, activitatea vagală ar putea crește, ceea ce duce la o FC mai mică (RRI mai mare). Se pare că nu numai activarea corticală induce o creștere aproape simultană a RRI (Barry, 1983; Damen și Brunia, 1987; Pfurtscheller și Lopes da Silva, 1999; Pfurtscheller et al., 2013), dar studiul nostru oferă primele dovezi că și un stimulator cardiac central din cerebel/trunchiul cerebral poate acționa ca forță motrice pentru oscilațiile RRI intrinseci și valurile spontane de respirație lentă. Această constatare susține cu tărie lucrările lui Perlitz et al. (2004) privind „ritmul de 0,15-Hz.”
Limitări și perspective viitoare
Selecția ROI-urilor cu semnale BOLD de bună calitate în cerebel/cefalus a fost făcută prin inspecție vizuală. Aceasta a inclus diferențierea între oscilațiile BOLD de origine neuronală și artefactele de mișcare. Acestea din urmă denotă semnale BOLD sincronizate în timp cu respirația. Pentru studii ulterioare, se recomandă calcularea valorilor de blocare a fazei (PLV; Pfurtscheller et al., 2017) între RRI și semnalele BOLD individuale în cerebel / trunchiul cerebral. În plus, evaluarea sincronă a metricilor de ventilație, cum ar fi volumul curent/volumul respirator pe timp, pCO2 la finalul respirației și tipul de respirație (nas vs. gură), precum și utilizarea diferitelor metode de denoising în contexte clinice și experimentale pe animale, ar putea oferi o perspectivă mai profundă asupra interacțiunilor complexe ale sistemelor de reglementare implicate.
„Deconectarea” aritmiei sinusale respiratorii este o excepție de la un fenomen fiziologic fundamental care poate apărea, de exemplu, în situații care provoacă anxietate. Dintre cei 23 de participanți sănătoși la un studiu fMRI, doar cinci subiecți au prezentat acest model de cuplaj paradoxal.
O condiție prealabilă pentru analiza BOLD (Bn și Bv) în trunchiul cerebral este o coincidență a respirației lente și a oscilațiilor RRI; cu toate acestea, un astfel de cuplaj 1:1 poate fi găsit doar la o minoritate de participanți. Prin urmare, acest tip de analiză este limitat la aceste cazuri rare. Cu toate acestea, am presupune că un stimulator cardiac central în trunchiul cerebral este, de asemenea, prevalent la persoanele cu o RSA normală și cuplaj 1:2 sau 1:3 (două sau trei respirații în timpul unui ciclu RRI). Primele rezultate din calculul cuplării de fază (PLV) între oscilațiile BOLD din trunchiul cerebral și oscilațiile RRI în banda de 0,1 până la 0,15-Hz susțin această presupunere. Cu această metodă, este posibil să se măsoare activitatea pacemakerului din trunchiul cerebral independent de ritmul de respirație. Această lucrare este în curs de desfășurare.
Trebuie remarcat faptul că nivelul anxietății de stat la indivizii cu respirație spontană lentă a variat între AS = 14 și AS = 28 (intervalul posibil al scorurilor AS: 10-40) și nu toți indivizii cu scoruri ridicate de anxietate au prezentat acest tip de respirație lentă. Prin urmare, concluzionăm că nu există o relație clară între respirația spontană lentă și procesarea anxietății.
Concluzie
1. Oscilațiile BOLD în stare de repaus de la nivelul cerebelului / trunchiului cerebral pot avea frecvențe alternante între 0,1 și 0,15 Hz, similare cu cele raportate în semnalele de RRI și respirație (Rassler et al., 2018).
2. Unele semnale BOLD de la nivelul cerebelului / trunchiului cerebral pot fi influențate de vasomoție (artera bazilară), în timp ce altele pot fi asociate cu o activitate de pacemaker central în trunchiul cerebral. În consecință, momentul real al începerii respirației, precum și sursa sa neuronală ar putea fi detectate în semnalele BOLD.
3. Ambele semnale BOLD cu origini diferite înregistrate simultan caracterizează o sursă comună, ceea ce susține lucrarea lui Perlitz et al. (2004).
4. Rezultatele sugerează că există dovezi pentru o respirație emoțională inconștientă la 6-9/min (0,1-0,15 Hz), care – într-un mod similar cu respirația conștientă de rezonanță la 6/min (0,1-0,15 Hz), care – într-un mod similar cu respirația conștientă de rezonanță la 6/min (0,1-0,15 Hz).1 Hz) care promovează bunăstarea psihologică (Mather și Thayer, 2018) – ridică, de asemenea, HRV și poate facilita procesarea emoțiilor negative.
Disponibilitatea datelor
Seturile de date generate pentru acest studiu sunt disponibile la cerere la autorul corespondent.
Declarație etică
Acest studiu a fost realizat în conformitate cu recomandările Declarației de la Helsinki din 1964, cu consimțământul informat în scris al tuturor subiecților. Toți subiecții și-au dat consimțământul informat în scris în conformitate cu Declarația de la Helsinki. Protocolul a fost aprobat de Comitetul de Etică de la Universitatea din Graz.
Contribuții ale autorilor
GP a contribuit la conceptualizare și la proiectul original. BR și AA au ajutat la metodologie, procesarea datelor, statistici, scriere și vizualizare. AS, BR, GS, GS, WK și JT au revizuit și editat manuscrisul.
Declarație privind conflictul de interese
Autorii declară că cercetarea a fost efectuată în absența oricăror relații comerciale sau financiare care ar putea fi interpretate ca un potențial conflict de interese.
Recunoștințe
Autorii doresc să le mulțumească lui Thomas Zussner, David Fink și Karl Koschutnig, Universitatea din Graz, pentru sprijinul acordat în achiziționarea datelor.
Abbreviații
BOLD, Dependent de nivelul de oxigenare a sângelui; ECG, Electrocardiogramă; (f)MRI, Imagistică prin rezonanță magnetică (funcțională); HR, Ritmul cardiac; HRV, Variabilitatea ritmului cardiac; PFC, Cortexul prefrontal; ROI, Regiunea de interes; RRI, Interval de bătaie la bătaie; RSA, Aritmie sinusală respiratorie.
Barry, R. J. (1983). Bradicardia primară și răspunsul cardiac evocat în contextul blocului operator. Physiol. Psychol. 11, 135-140. doi: 10.3758/BF03326784
CrossRef Full Text | Google Scholar
Birn, R. M., Diamond, J. B., J. B., Smith, M. A., and Bandettini, P. A. (2006). Separarea fluctuațiilor legate de variația respiratorie de fluctuațiile legate de activitatea neuronală în fMRI. NeuroImage 31, 1536-1548. doi: 10.1016/j.neuroimage.2006.02.048
PubMed Abstract | CrossRef Full Text | Google Scholar
Buxton, R. B., Uludag, K., Dubowitz, D. J., și Liu, T. T. (2004). Modelarea răspunsului hemodinamic la activarea creierului. NeuroImage 23, 220-233. doi: 10.1016/j.neuroimage.2004.07.013
CrossRef Full Text | Google Scholar
Damen, E. J. P., and Brunia, C. H. M. (1987). Modificări ale frecvenței cardiace și ale potențialelor cerebrale lente legate de pregătirea motorie și anticiparea stimulilor într-o sarcină de estimare a timpului. Psychophysiology 24, 700-713. doi: 10.1111/j.1469-8986.1987.tb00353.x
PubMed Abstract | CrossRef Full Text | Google Scholar
Eckberg, D. L. (2003). Poarta respiratorie umană. J. Physiol. 548, 339-352. doi: 10.1113/jphysiol.2002.037192
PubMed Abstract | CrossRef Full Text | Google Scholar
Häbler, H.-J., and Jänig, W. (1995). Coordonarea sistemelor simpatic și respirator: experimente neurofiziologice. Clin. Exp. Hypertens. 17, 223-235. doi: 10.3109/10641969509087067
CrossRef Full Text | Google Scholar
Haselton, J. R., și Guyenet, P. G. (1989). Modularea respiratorie centrală a neuronilor simpaticoexcitatori medulari la șobolan. Am. J. Phys. 256, R739-R750. doi: 10.1152/ajpregu.1989.256.3.R739
PubMed Abstract | CrossRef Full Text | Google Scholar
Homma, I., and Masaoka, Y. (2008). Ritmurile de respirație și emoțiile. Exp. Physiol. 93, 1011-1021. doi: 10.1113/expphysiol.2008.042424
PubMed Abstract | CrossRef Full Text | Google Scholar
Julien, C. (2006). Enigma undelor Mayer: fapte și modele. Cardiovasc. Res. 70, 12-21. doi: 10.1016/j.cardiores.2005.11.008
PubMed Abstract | Full CrossRef Full Text | Google Scholar
Kato, A., Takahashi, K., și Homma, I. (2018). Relațiile dintre trăsăturile și parametrii respiratori în timpul respirației liniștite la subiecții normali. J. Physiol. Sci. 68, 369-376. doi: 10.1007/s12576-017-0539-7
PubMed Abstract | CrossRef Full Text | Google Scholar
Lambertz, M., and Langhorst, P. (1998). Modificări simultane ale organizării ritmice în neuronii trunchiului cerebral, respirația, sistemul cardiovascular și EEG între 0,05 Hz și 0,5 Hz. J. Auton. Nerv. Syst. 68, 58-77.
Google Scholar
Mandel, D. A., și Schreihofer, A. M. (2006). Modularea respiratorie centrală a neuronilor barosensibili în medulla ventrolaterală ventrolaterală caudală de șobolan. J. Physiol. 572, 881-896. doi: 10.1113/jphysiol.2005.103622
PubMed Abstract | CrossRef Full Text | Google Scholar
Mateo, C., Knutsen, P. M., Tsai, P. S., Shih, A. Y., și Kleinfeld, D. (2017). Intrarea fluctuațiilor vasomotorii ale arteriolelor de către activitatea neuronală este o bază a conectivității „în stare de repaus” dependentă de nivelul de oxigenare a sângelui. Neuron 96, 1-13. doi: 10.1016/j.neuron.2017.10.012
CrossRef Full Text | Google Scholar
Mather, M., and Thayer, J. (2018). Cum afectează variabilitatea ritmului cardiac rețelele cerebrale de reglementare a emoțiilor. Curr. Opin. Behav. Sci. 19, 98-104. doi: 10.1016/j.cobeha.2017.12.017
PubMed Abstract | CrossRef Full Text | Google Scholar
Moeller, S., Yacoub, E., Olman, C. A., Auerbach, E., Strupp, J., Harel, N., et al. (2010). GE-EPI multibandă multislice GE-EPI la 7 Tesla, cu o accelerare de 16 ori folosind imagistica paralelă parțială cu aplicare la fMRI spațial și temporal ridicat la nivelul întregului creier. Magn. Reson. Med. 63, 1144-1153. doi: 10.1002/mrm.22361
PubMed Abstract | Full Text | Google Scholar
Munn, Z., Moola, S., Lisy, K., Riitano, D., și Murphy, F. (2015). Claustrofobia în imagistica prin rezonanță magnetică: o revizuire sistematică. Radiography 21, e59-e63. doi: 10.1016/j.radi.2014.12.004
CrossRef Full Text | Google Scholar
Obrig, H., Neufang, M., Wenzel, R., Kohl, M., Steinbrink, J., Einhäupl, K., et al. (2000). Oscilații spontane de frecvență joasă ale hemodinamicii și metabolismului ccerebral la adulții umani. NeuroImage 12, 623-639. doi: 10.1006/nimg.2000.0657
PubMed Abstract | CrossRef Full Text | Google Scholar
Perlitz, V., Lambertz, M., Cotuk, B., Grebe, R., Vandenhouten, R., Flatten, G., et al. (2004). Ritmurile cardiovasculare în banda de 0,15-Hz: originea comună a unor fenomene identice la om și câine în formațiunea reticulară a trunchiului cerebral? Eur. J. Phys. 448, 579-591. doi: 10.1007/s00424-004-1291-4
PubMed Abstract | CrossRef Full Text | Google Scholar
Pfurtscheller, G., și Lopes da Silva, F. (1999). Sincronizarea și desincronizarea EEG/MEG legate de evenimente: principii de bază. Clin. Neurophysiol. 110, 1842-1857. doi: 10.1016/S1388-2457(99)00141-8
PubMed Abstract | Refef Full Text | Google Scholar
Pfurtscheller, G., Schwerdtfeger, A., Seither-Preisler, A., Brunner, C., Aigner, C. S., Brito, J., et al. (2017). Comunicarea creier-inimă: dovezi pentru oscilațiile „pacemakerului central” cu o frecvență dominantă la 0,1 Hz în cingulum. Clin. Neurophysiol. 128, 183-193. doi: 10.1016/j.clinph.2016.10.097
PubMed Abstract | CrossRef Full Text | Google Scholar
Pfurtscheller, G., Schwerdtfeger, A., Schwerdtfeger, A., Seither-Preisler, A., Brunner, C., Aigner, C. S., Calisto, J., et al. (2018). Sincronizarea oscilațiilor intrinseci de 0,1-Hz dependente de nivelul de oxigen din sânge în amigdala și cortexul prefrontal la subiecții cu anxietate de stare crescută. Eur. J. Neurosci. 47, 417-426. doi: 10.1111/ejn.13845
PubMed Abstract | Ref. Full Text | Google Scholar
Pfurtscheller, G., Solis-Escalante, T., Barry, R. J., Klobassa, D. S., Neuper, C. și Müller-Putz, G. R. (2013). Frecvența cardiacă rapidă și modificările EEG în timpul executării și reținerii imaginilor motorii de picior cu ritm de cue. Front. Hum. Neurosci. 7:379. doi: 10.3389/fnhum.2013.00379
PubMed Abstract | Cross Full Text | Google Scholar
Rassler, B., Schwerdtfeger, A., Aigner, C. S., și Pfurtscheller, G. (2018). „Deconectarea” aritmiei sinusale respiratorii poate apărea la o minoritate de subiecți în timpul imagisticii prin rezonanță magnetică funcțională (fMRI). Front. Physiol. 9: 1688. doi: 10.3389/fphys.2018.01688
PubMed Abstract | CrossRef Full Text | Google Scholar
Thayer, J. F., și Lane, R. D. (2009). Claude Bernard și conexiunea inimă-creier: elaborarea ulterioară a unui model de integrare neuroviscerală. Neurosci. Biobehav. Rev. 33, 81-88. doi: 10.1016/j.neubiorev.2008.08.004
PubMed Abstract | CrossRef Full Text | Google Scholar
Tzourio-Mazoyer, N., Landeau, B., Papathanassiou, D., Crivello, F., Etard, O., Delcroix, N., et al. (2002). Etichetarea anatomică automatizată a activărilor în SPM folosind o parcelare anatomică macroscopică a creierului MNI MRI cu un singur subiect. NeuroImage 15, 273-289. doi: 10.1006/nimg.2001.0978
PubMed Abstract | Full CrossRef Text | Google Scholar
Yasuma, F., and Hayano, J. (2004). Aritmia sinusală respiratorie: de ce se sincronizează bătăile inimii cu ritmul respirator? Chest 125, 683-690. doi: 10.1378/chest.125.2.683
PubMed Abstract | Refef Full Text | Google Scholar
Zaccaro, A., Piarulli, A., Laurino, M., Garbella, E., Menicucci, D., Neri, B., et al. (2018). Cum controlul respirației vă poate schimba viața: o revizuire sistematică a corelațiilor psiho-fiziologice ale respirației lente. Front. Hum. Neurosci. 12:353. doi: 10.3389/fnhum.2018.00353. eCollection 2018.
CrossRef Full Text | Google Scholar
.