ADVERTISMENTE:
În acest articol vom discuta despre Paramecium Caudatum: – 1. Habitatul, habitatul și cultura Paramecium Caudatum 2. Habitatul, habitatul și cultura Paramecium Caudatum. Structura lui Paramecium Caudatum 3. Locomoția 4. Nutriție 5. Respirație și excreție 6. Osmoreglarea 7. Comportamentul 8. Reproducerea 9. Comportamente aberante în timpul reproducerii 10. Unele particule citoplasmatice.
Contenit:
- Habitat, Habitatul și cultura Paramecium Caudatum
- Structura Paramecium Caudatum
- Locomoția Paramecium Caudatum
- Nutriția la Paramecium Caudatum
- Respirația și excreția la Paramecium Caudatum
- Osmoreglarea Paramecium Caudatum
- Comportamentul lui Paramecium Caudatum
- Reproducerea la Paramecium Caudatum
- Comportamentul aberant în reproducere la Paramecium Caudatum
- Câteva particule citoplasmatice semnalate la Paramecium Caudatum
. Paramecium Caudatum
- Habitat, Habitat și cultură de Paramecium Caudatum:
- Cultura de Paramecium:
- Structura Paramecium caudatum:
- (i) Mărimea și forma Paramecium caudatum:
- (ii) Peliculă:
- (iii) Cili:
- (iv) Sistemul infra-ciliar:
- (v) Canelura orală și citotipul:
- (vi) Citoplasma:
- (vii) Ectoplasma:
- (viii) Trichocistele:
- (ix) Neuromotoriul și fibrilele asociate:
- (x) Endoplasma:
- (xi) Vacuole contractile:
- (xii) Vacuole alimentare:
- Locomoția lui Paramecium Caudatum:
- (i) Metabolia sau contorsionarea corpului:
- (ii) Locomoția ciliară:
- Nutriția lui Paramecium Caudatum:
- Mecanism de hrănire:
- Digestia și digestia la Paramecium Caudatum:
- Respirația și excreția la Paramecium Caudatum:
- Comportamentul lui Paramecium Caudatum:
- (i) Reacții la contact (Thigmotaxie):
- (ii) Reacții la substanțe chimice (chemotaxie):
- (iii) Reacții la temperatură (termotaxie):
- (iv) Reacții la lumină (fototaxie):
- (v) Reacții la curentul electric (galvanotaxie):
- (vi) Reacții la curentul de apă (Rheotaxis):
- (vii) Reacții la gravitație (Geotaxie):
- Reproducere la Paramecium Caudatam:
- (i) Fisiunea binară transversală:
- (ii) Conjugarea:
- Comportamentul aberant în reproducere la Paramecium Caudatum:
- (i) Endomixia:
- (ii) Autogamia:
- (iii) Hemixis:
- (iv) Citogamia:
- Câteva particule citoplasmatice raportate la Paramecium Caudatum:
- (i) Particule Kappa:
- (ii) mµ, Particule:
- (iii) Particulele Pi:
- (iv) Particulele Lambda:
Habitat, Habitat și cultură de Paramecium Caudatum:
Paramecium caudatum (Gr., paramekes = alungit; L., caudata = coadă) se găsește în mod obișnuit în iazuri de apă dulce, bălți, șanțuri, cursuri de apă, lacuri, rezervoare și râuri. Se găsește în mod special în abundență în iazurile stagnante bogate în materii în descompunere, în infuzii organice și în apele de canalizare. Paramecium caudatum este un organism care trăiește liber și această specie este răspândită în întreaga lume.
Cultura de Paramecium:
ADVERTISMENTE:
Se iau buruieni scufundate dintr-un iaz și se pun într-un borcan cu apă distilată, se acoperă borcanul și se lasă să putrezească; în câteva zile vor apărea roiuri de Paramecia. Acum fierbeți fânul în apă, decantați infuzia, adăugați câteva boabe de grâu și lăsați-o să stea până când se tulbură cu bacterii.
Transferați Paramecia din primul borcan în acest lichid, unde se vor înmulți rapid. Infuziile de fân singure vor produce Paramecia care arată prezența chisturilor, și au fost raportate chisturi asemănătoare cu boabe de nisip, dar nu există nici o dovadă că Paramecia formează chisturi, deoarece acestea nu au fost niciodată confirmate.
Structura Paramecium caudatum:
(i) Mărimea și forma Paramecium caudatum:
Paramecium caudatum (Fig. 20.1) este un organism microscopic și este vizibil cu ochiul liber ca un corp alungit minuscul.
ADVERTISMENTE:
Apare de culoare gri deschis sau alb măsurând în mod obișnuit între 170 și 290 microni în lungime și poate atinge o lungime de până la 300-350 microni. P. caudatum seamănă cu talpa unui papuc sau a unui pantof, prin urmare, animalul este cunoscut în mod obișnuit sub numele de animalcul papuc. Are o lungime de patru ori mai mare decât lățimea și este oarecum cilindrică, cu capete net diferite.
Partea anterioară care se deplasează înainte este subțire, cu un capăt bont sau rotunjit, în timp ce capătul posterior este oarecum ascuțit sau în formă de con. Cea mai lată parte a organismului se află chiar sub mijloc. Corpul animalului are o formă asimetrică, prezentând o suprafață orală sau ventrală bine definită și una abdominală sau dorsală.
(ii) Peliculă:
Corpul este acoperit de o peliculă subțire, dublu stratificată, elastică și fermă, formată din gelatină. Pelicula menține forma animalului, dar este suficient de elastică pentru a permite contracții. Pelicula are o membrană dublă, membrana exterioară este continuă cu cilia, iar cea interioară cu ectoplasma. La o mărire mai mare a microscopului, pelicula prezintă pe suprafața sa depresiuni dreptunghiulare sau hexagonale.

Acest aranjament se păstrează pe suprafața dorsală a Paramecium, dar pe suprafața ventrală crestele converg în față și în spate spre o deschidere preorală și postorală. Fiecare depresiune hexagonală este perforată de o deschidere centrală prin care iese un singur ciliu. Marginile anterioare și posterioare ale depresiunilor hexagonale poartă deschiderile trichocisturilor.
Studiul la microscopul electronic al peliculei (Fig. 20.2) realizat de Ehret și Powers (1957) a arătat că depresiunile hexagonale corespund unor serii regulate de cavități, alveolele. Toți alveolele formează în mod colectiv un strat alveolar continuu, care este delimitat de o membrană alveolară externă și una internă.
Stratul extern se află în contact strâns sub membrana celulară externă. Prin urmare, pelicula include membrana celulară externă, membrana alveolară externă și membrana alveolară internă.
(iii) Cili:
Întregul corp este acoperit cu numeroase proiecții mici, asemănătoare unor fire de păr, numite cili. Cilii apar în rânduri longitudinale pe tot corpul, această stare este cunoscută sub numele de holotricot, în care cilia corpului sunt egali. Cilii au aceeași structură ca și flagelii, au un înveliș protoplasmatic extern sau o membrană plasmatică cu nouă fibrile longitudinale duble într-un inel periferic. La unii cili, cele nouă fibrile exterioare nu sunt împerecheate.
ADVERTISMENTE:
Există două fibrile longitudinale centrale care sunt mai subțiri decât fibrilele exterioare. Fiecare ciliu provine dintr-un granule bazal sau kinetosom. Cele nouă perechi de fibrile periferice fuzionează împreună pentru a forma peretele kinetosomului, astfel, kinetosomul este un tub care este fie deschis, fie închis la capătul său inferior, cele două fibrile centrale se opresc la nivelul peliculei la majoritatea ciliatelor.
Din cinetosom se desprinde un rizoplast subțire care nu se unește cu nucleul. Multe metazoare au, de asemenea, cili, structura lor este aceeași, cu excepția faptului că granulația bazală este diferită și are filamente fine sau fibre de înrădăcinare care se întind în citoplasmă. Dar cilia diferă de flageli prin faptul că sunt, în general, mai numeroși și de dimensiuni mai mici.
Ciliatura poate fi împărțită convenabil în cili corporali sau somatici, care se găsesc pe suprafața corpului, și în ciliatură orală, care este asociată cu regiunea gurii. Cilii corpului sunt egali, dar sunt mai lungi la capătul posterior, de unde și denumirea de caudatum. Ciliile sunt organite de locomoție și de colectare a hranei, ele acționează, de asemenea, ca receptori senzoriali și detectează stimulii din mediul extern.

Ultra structură a cililor:
Cilii și flagelii au o compoziție fibrilară. La bază, ciliul are diametrul de aproximativ 0,2 microni sau 2.000 A0 care poate fi până la 10 microni deasupra suprafeței celulare. Cilii sunt delimitați de o membrană unitară cu o grosime de 90 A0 care se aseamănă și rămâne continuă cu membrana plasmatică. Spațiul delimitat al ciliului conține o substanță apoasă cunoscută sub numele de matrice.
În matrice, rămân înglobate unsprezece fibrile longitudinale sau microtubuli. Dintre cele unsprezece fibrile, două sunt situate în centru, în timp ce restul de nouă fibrile rămân dispuse periferic în jurul fibrilelor centrale. Fiecare dintre cele nouă fibrile exterioare are un diametru de 360A0 și este compusă din două sub-fibrile cu diametrul cuprins între 180 și 250A0.
Aceste sub-fibrile sunt desemnate ca fiind sub-fibrila A și sub-fibrila B.
ADVERTISMENTE:
Sub-fibrila A este puțin mai mare decât sub-fibrila B. Sub-fibrila A dă două proeminențe groase sau brațe de o parte a sa. Brațele sub-fibrilei A din toate fibrilele exterioare rămân orientate în sensul acelor de ceasornic. În plus, subfibrila A se află mai aproape de centrul ciliului decât subfibrila B. Ambele subfibrile au un perete comun cu o grosime de 50A0.
Cele două fibrile centrale nu au sub-fibrilele împerecheate ca cele nouă fibrile periferice, ci fiecare conține doar un singur tubul. Fiecare fibulă centrală are un diametru de aproximativ 250A0 și este compusă dintr-un perete de 60A0 grosime.
Ambele fibrile centrale rămân separate de un spațiu de 350A0 și rămân învelite într-un înveliș comun. Gibbnos (1967) a raportat că învelișul fibrilelor centrale dă nouă legături sau raze orientate radial pentru fiecare sub-fibrilă A.

Microscopia electronică de înaltă rezoluție a arătat că fiecare dintre fibrilele periferice și centrale ale ciliilor și flagelilor este compusă din zece până la douăsprezece filamente de 40A0 grosime. Fiecare filament este mărginit. Fiecare fagure rămâne dispus în rețele de 40 pe 50A0 în planul peretelui tubului. Aceste mărgele sunt considerate ca fiind subunitatea de bază a structurii tubului.
(iv) Sistemul infra-ciliar:
Sistemul infraciliar este situat chiar sub alveolele peliculare. Este format din kinetosom sau corpul bazal și kinetodesma. Cilii iau naștere din kinetosomi și din fiecare kinetosom ia naștere o fibrilă citoplasmatică delicată numită kinetodesmă, (Fig. 20.2).
Pe sub peliculă, ușor spre dreapta, dar unită cu toți kinetozomii dintr-un singur rând longitudinal, se află un fascicul longitudinal de mai multe kinetodesme, o kinetodesmă a fiecărui kinetozom se prelungește pe o distanță anterioară în propriul său fascicul de kinetodesme.
Un rând longitudinal de kinetozomi cu kinetodesmele lor formează o unitate longitudinală numită kinetodezom. Toate kinețiile sau kinetia alcătuiesc sistemul infraciliar al unui ciliat. Kinetia se află în cortex, sub peliculă, iar numărul lor este destul de constant pentru fiecare ciliat.
Sistemul infraciliar controlează și coordonează mișcările ciliatelor și determină formarea organitelor în diviziunea celulară, de exemplu, unele kineții formează gura. În fisiunea binară a ciliatelor, kinetia este tăiată transversal în două, fiecare mergând la o celulă fiică, aceasta se numește fisiune perikinetală.
(v) Canelura orală și citotipul:
Pe partea ventrolaterală se află o depresiune mare, oblică și puțin adâncă, numită șanț oral sau peristom, care conferă animalului un aspect asimetric. Acesta se întinde oblic spre înapoi dintr-o parte (de obicei de la stânga la dreapta, dar în unele cazuri de la dreapta la stânga) și se termină puțin în spatele corpului mijlociu. Canelura bucală duce la o scurtă depresiune conică în formă de pâlnie numită vestibul.
ADVERTISMENTE:
Vestibulul duce direct în deschiderea fixă, de formă ovală, numită citostom (gură). Extinzându-se direct de la citostom spre centrul corpului se află citofaringele larg. Citofaringele se întoarce apoi brusc spre partea posterioară pentru a deveni esofagul subțire și conic.
Astfel, esofagul este aproximativ paralel cu suprafața corpului lui Paramecium, cu excepția extremității sale posterioare. Aici, esofagul se întoarce din nou spre centrul animalului pentru a duce în vacuola alimentară în formare.
Citotipul (denumit și anus celular sau punct anal sau citoproct) se află pe suprafața ventrală a corpului, aproape vertical, în spatele citostomului sau gurii. Particulele de hrană nedigerate sunt eliminate prin citotige. Cilarea citofaringelui este foarte complicată.
Gelei (1934) a raportat prezența a patru rânduri, iar Lund (1941) a observat cel puțin patru rânduri. O structură numită penniculus se găsește pe peretele stâng al citofaringelui și se rotește în spirală la aproximativ 90 de grade, astfel încât extremitatea sa posterioară se află pe suprafața orală (ventrală) a esofagului.
Potrivit lui Lund, penniculul este format din opt rânduri de cili dispuși în două blocuri apropiate de fiecare. O bandă similară compusă din patru rânduri de cili lungi care sunt mai puțin compacți decât în penniculus este denumită quadrulus. Aceasta coboară în spirală pe peretele dorsal al cavității bucale și se termină aproape de penniculus.
Peniculul și cvadrulul au fost numite în mod greșit de unii lucrători o membrană ondulată. Quadrulus și penniculus controlează trecerea alimentelor. Nu se știe cum funcționează cilia, probabil că fibrilele lor se contractă în mod ritmic, ceea ce determină îndoirea. Gelei (1925) a precizat că funcția penniculului este forțarea elementelor alimentare în organism.

(vi) Citoplasma:
Citoplasma se diferențiază într-o zonă îngustă, externă sau corticală, numită ectoplasmă și o regiune mai mare, internă sau medulară, numită endoplasmă.
(vii) Ectoplasma:
Ectoplasma (ectosarc sau cortex) este o parte permanentă a corpului, delimitată în mod frapant de endoplasmă. Ectoplasma formează un strat exterior ferm, clar, subțire și dens. Acesta conține tricocite, cili și structuri fibrilare și este delimitat la exterior de un înveliș numit peliculă.
(viii) Trichocistele:
Încrustate în ectoplasmă, în unghiuri drepte față de suprafață, se află mici pungi fusiforme numite trichociste. Un punct mic pe fiecare margine anterioară și posterioară a hexagonului marchează poziția unui tricocist. Aceștia sunt umpluți cu un fluid refractat, dens, având o substanță umflătoare, la capătul exterior se află un cap conic sau un spike.
Tricocistele se află perpendicular în ectoplasmă, se deschid prin pori mici pe crestele zonelor hexagonale ale peliculei.
Se nasc din kinetosomii de cili, apoi migrează și se localizează la distanță egală în endoplasmă. Când animalul este iritat, tricocitele sunt evacuate sub formă de fire lungi și lipicioase. Un trichocist descărcat are un vârf opac, asemănător unui cui inversat, și un arbore lung și striat, dar arborele nu se vede în stare nedeversată și se formează probabil în timpul descărcării.
Funcția trichocisturilor este incertă, dar ele sunt descărcate ca reacție la contacte locale și leziuni, ele pot servi ca organite de apărare.
Dar acest lucru este incert, deoarece trichocistele sunt ineficiente împotriva Didinium, principalul prădător al Paramecium, ele pot servi la fixarea animalului într-un loc în timpul hrănirii. La unele ciliate, trichocistele acționează ca organite de ofensă. După ce tricocistele sunt evacuate, regenerate din kinetosomi.


(ix) Neuromotoriul și fibrilele asociate:
Potrivit lui Lund (1933), pe peretele dorsal stâng al citofaringelui, aproximativ la nivelul marginii posterioare a citostomului, se află o masă foarte mică, bilobată, neuromotoriul. Din neuromotorium, fibrilele radiază în endoplasmă.
Dintre acestea, patru sau mai multe trec de obicei aproape de peretele dorsal al corpului, dar restul sunt mai scurte și nu au o poziție precisă. Toate sunt denumite fibrile endoplasmatice. Funcția lor este necunoscută, dar este posibil să coordoneze mișcările de hrănire ale ciliilor bucali. De asemenea, fibrilele pot oferi suport mecanic, elasticitate, contractilitate, conductivitate și influență metabolică.

(x) Endoplasma:
Edoplasma sau medula este partea mai fluidă și mai voluminoasă a citoplasmei care conține multe granule citoplasmatice, precum și alte incluziuni și structuri de natură specializată. Incluziunile citoplasmatice sunt mitocondriile, aparatele Golgi, vacuolele, cristalele, granulele și cromidele etc. În endoplasmă se mai găsesc și alte structuri, și anume nuclei, vacuole contractile și vacuole alimentare.
Nuclee:
În endoplasmă, în apropierea citostomului, se găsesc două nuclee, adică Paramecium este heterocariot, un macronucleu mare elipsoidal și granular și un alt micronucleu mic și compact.
Macronucleul este un corp vizibil, elipsoidal sau în formă de rinichi. Este de tip compact care conține fire fine și granule de cromatină discrete, strâns compacte, de dimensiuni variabile și înglobate într-o matrice acromatică. Acesta posedă mulți nucleoli și mult mai mult material cromatinic (ADN).
Este nucleu somatic sau vegetativ. Se divide amitotic și controlează funcțiile vegetative (activități metabolice) ale animalului. Suferă mitoză.
Micronucleul este mic, compact și sferic. Se găsește, în general, în apropierea macronucleului, adesea într-o concavitate. Granulele și firele fine de cromatină sunt distribuite uniform în întreaga structură. Micronucleul se divide mitotic și controlează reproducerea.
Numărul de micronucleu variază în funcție de specie; este unul la P. caudatum, două la P. aurelia și multe la P. multimicronucleatum. Micronucleul conține un nucleolus distinct la P. aurelia, dar nu se găsește la P. caudatum. Moses (1949; 1950) a raportat că macronucleul și micronucleul au o compoziție chimică identică.
(xi) Vacuole contractile:
Există două vacuole contractile mari, pline de lichid, fiecare situată în apropierea unui capăt al corpului, aproape de suprafața dorsală. Poziția lor este fixă (spre deosebire de Amoeba), ele se află între ectoplasmă și endoplasmă, dar sunt organite temporare care dispar periodic. La unele specii, ele par să aibă o membrană de căptușeală, caz în care nu dispar în întregime în timpul sistolei.
Conectate la fiecare vacuolă contractilă sunt cinci până la douăsprezece canale radiante tubulare, fiecare constând dintr-o parte terminală, o ampulă lungă care se prăbușește când este goală și un canal injector scurt care se deschide în vacuolă.
Paramecium. Canal radial. A – gol; B – plin.
Canalele comunică cu o mare parte a corpului de unde preiau lichidele și le toarnă în vacuolă care, astfel, se reconstituie și crește în dimensiuni, când vacuola contractilă ajunge la dimensiunea maximă se contractă brusc (sistole) și își evacuează conținutul printr-un por permanent din peliculă, apoi canalele formează din nou vacuolele contractile, canalele nu dispar în întregime, deoarece sunt structuri permanente.
Cele două vacuole contractile se descarcă neregulat, cea posterioară se contractă mai repede pentru că este aproape de citofaringe și în ea intră mai multă apă. Funcția principală a canalelor și a vacuolelor contractile este hidrostatică, ele elimină excesul de apă din protoplasmă, apa este parțial absorbită și parțial absorbită în timpul hrănirii.
Substanța azotată reziduală are compuși de amoniac și unii urați care sunt expulzați din vacuolele contractile împreună cu CO2, dar nu există nici o dovadă că materia excretoare este secretată de protoplasmă în canale sau vacuole contractile.
Există un citotip sau anus permanent fix sub peristomul de o parte și de alta a citofaringelui, prin care sunt expulzate resturile nedigerate de hrană și urații.
Studiul la microscopul electronic al vacuolelor contractile a evidențiat faptul că fiecare aparat contractil este format din o parte din tubulii reticulului endoplasmatic, tubulii nefriciali, canalele de alimentare, vacuolele accesorii (canalele radiale) și vacuola contractilă principală. Se presupune că vacuolele accesorii sunt ampule ale canalelor de alimentare (Fig. 20.11).
(xii) Vacuole alimentare:
Sunt corpuri aproximativ sferice, necontractile, care variază ca mărime și număr, situate în endoplasmă. Ele conțin particule de hrană ingerată, în principal bacterii și o cantitate mică de lichid delimitată de o membrană definită subțire. Volkonsky (1934) a propus denumirea de gastrioli pentru aceste vacuole. Asociați cu vacuolele alimentare sunt granulele digestive.
Locomoția lui Paramecium Caudatum:
Paramecium Caudatum se deplasează prin două metode, și anume, prin metabolism sau contorsionări ale corpului și prin cili.
(i) Metabolia sau contorsionarea corpului:
Corpul lui Paramecium Caudatum posedă elasticitate, se poate strecura printr-un pasaj mai îngust decât corpul său, după care corpul își reia forma normală. Această schimbare temporară a formei corpului este metabolie, ea este provocată la Paramecium de către protoplasmă.
(ii) Locomoția ciliară:
Locomoția determinată de cili este principala metodă. Cilii pot bate înainte sau înapoi, permițând animalului să înoate anterior sau posterior.
În mod normal, animalul înoată înainte, cilia bătând în spate, dar oblic, cilia se rigidizează și se îndoaie rapid în spate până aproape atinge suprafața corpului, aceasta se numește cursă efectivă; apoi cilia devin moi și revin încet la poziția verticală inițială, aceasta este , numită cursă de recuperare.
Cilii din același rând transversal bat împreună, iar cei din același rând longitudinal bat unul după altul, de la capătul anterior la cel posterior.

Această mișcare coordonată a ciliilor se numește ritm metacronic, care se datorează sistemului infraciliar; aceasta determină înotul înainte al animalului. Dar atunci când cilia corpului bate oblic spre înapoi, atunci, în același timp, cilia mai lungă a șanțului oral bate mai viguros, ceea ce face ca extremitatea anterioară să se abată spre stânga.
Acțiunea ciliilor corpului și ai șanțului bucal face ca animalul să se rotească pe axa sa lungă. Această rotație este întotdeauna spre stânga (cu excepția lui P. calkinsi care se rotește în spirală spre dreapta).
Această combinație de mișcare înainte, de rotire și rotație face ca animalul să se deplaseze înainte pe o traiectorie spiralată în sens invers acelor de ceasornic. Această traiectorie are o axă dreaptă, iar aceeași suprafață corporală a animalului rămâne spre axa traiectoriei în spirală. Însă, atunci când înoată în sens invers, toate speciile se rotesc spre dreapta.
Bătălia ciliară poate fi inversată astfel încât ciliile să se deplaseze oblic înainte prin care animalul înoată înapoi. Prin acțiunea ciliară, Paramecium se deplasează cu o viteză de 1500 microni sau chiar mai mult pe secundă.
Jennings susține că spiralarea lui Paramecium se datorează faptului că, în timp ce cilia lovește în principal în spate, o face oblic spre dreapta, determinând astfel animalul să se rostogolească spre stânga.
De asemenea, această rotire a corpului spre suprafața abdominală se datorează în mare parte puterii mai mari a loviturii efective a ciliilor orali care lovesc mai direct în spate. Rezultatul – rotația Parameciumului pe axa sa lungă – permite astfel Parameciumului să urmeze un curs mai mult sau mai puțin drept în formarea unor spirale mari.
Nutriția lui Paramecium Caudatum:
La Paramecium Caudatum, nutriția este holozoică. Hrana cuprinde în principal bacterii și protozoare minuscule. Paramecium nu așteaptă hrana, ci o vânează în mod activ.
Se susține că Paramecium Caudatum manifestă o alegere în selectarea hranei sale, dar nu pare să existe niciun temei pentru acest lucru, deși înghite doar anumite tipuri de bacterii; datele disponibile sugerează că între 2 și 5 milioane de indivizi de Bacillus coli sunt devorați de un singur Paramecium în 24 de ore. De asemenea, se hrănește cu plante unicelulare, cum ar fi algele, diatomeele etc., precum și cu bucăți mici de animale și legume.
Mecanism de hrănire:
Când Paramecium Caudatum intră într-o regiune cu hrană abundentă, vine să se odihnească. Se hrănește numai în repaus sau când înoată foarte încet, nu se hrănește niciodată când înoată repede. Bătaia ciliilor din șanțul bucal face ca un vârtej conic de apă încărcată cu hrană să fie măturat în șanțul bucal de la o distanță în avans față de capătul anterior (Fig. 20.16).
Particulele de hrană se îndreaptă apoi spre vestibulul de unde unele particule de hrană sunt respinse și aruncate afară, dar altele trec în citostom.
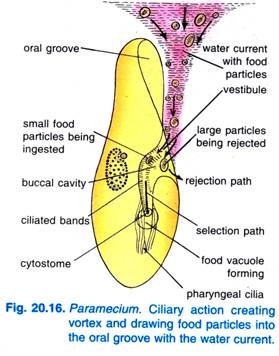
La capătul citofaringelui se formează un vacuol alimentar care se umple cu particule de hrană. Quadrulus și peniculi controlează trecerea alimentelor în vacuola alimentară care se formează lateral. Atunci când vacuola alimentară atinge o anumită dimensiune, fibrele post-bucale prind vacuola alimentară și aceasta este strânsă de ele și își continuă drumul.
Vacuolul conține, pe lângă hrană, și puțină apă. Mișcările rotative de curgere a endoplasmei, numite ciclosis, transportă vacuolele alimentare de-a lungul unui traseu definit care este echivalent din punct de vedere funcțional cu un tub digestiv.
Tractul începe de la capătul citofaringelui, apoi spre partea posterioară, apoi înainte pentru a circula împreună cu endoplasma, apoi spre suprafața dorsală, apoi spre capătul anterior, apoi în jos spre citoputere. La începutul călătoriei sale, vacuola alimentară scade în dimensiune, apoi crește din nou.

Digestia și digestia la Paramecium Caudatum:
În timpul ciclozei, digestia are loc prin enzimele secretate de protoplasmă în vacuole. În digestie, proteinele sunt transformate în aminoacizi, carbohidrații în zaharuri solubile și glicogen, iar grăsimile sunt, probabil, de asemenea, digerate.
Contenutul vacuolelor alimentare este la început acid (pH aproximativ 4) și apoi devine alcalin, digestia majoră are loc în timpul fazei alcaline. Materia nedigerată este eghițată prin citoputere cu o anumită forță.
Cicloza poate fi demonstrată experimental; dacă Parameciumului i se dă să mănânce lapte colorat cu roșu Congo, globulele de grăsime din lapte din vacuolele alimentare vor deveni mai întâi roșii datorită reacției acide a enzimelor, apoi vor trece de la nuanțe de violet la albastru datorită reacției alcaline, vacuolele vor arăta cursul ciclicității.
Respirația și excreția la Paramecium Caudatum:
Schimbul de gaze (oxigen și dioxid de carbon) are loc prin pelicula semipermeabilă, ca și la alți protozoare de apă dulce, prin procesul de difuzie. Paramecium Caudatum își obține oxigenul din apa înconjurătoare. Dioxidul de carbon și deșeurile organice, cum ar fi amoniacul, rezultate din metabolism, sunt probabil excretate prin difuzie în apă în sens invers.

6. Osmoreglarea la Paramecium Caudatum:
Paramecium Caudatum are doi vacuoli contractili, unul anterior și unul posterior. Funcția vacuolelor contractile este osmoreglarea, adică reglarea conținutului de apă din organism și poate servi, de asemenea, la excreția deșeurilor azotate, cum ar fi ureea și amoniacul.
Excesul de apă (din cauza endosmozei continue) din interiorul citoplasmei este secretat în tubulii reticulului endoplasmatic și se îndreaptă spre tubulii nefriciali → canalele de alimentare → și se colectează în ampulă dintr-o serie de 6 până la 11 canale radiante care converg spre și se descarcă în fiecare vacuolă. Canalele sunt mai vizibile în momentul în care se formează un vacuol.
Când fiecare vacuolă se umflă (diastole) până la o anumită dimensiune, se contractă (sistole) și se descarcă în exterior, probabil printr-un por. Vacuolele contractile se contractă alternativ, la intervale de 10 – 20 de secunde.
Vacuola contractilă posterioară lucrează mai repede decât vacuola anterioară din cauza admiterii unei cantități mari de apă în regiunea posterioară de către citofaringe. Vacuolele contractile mențin o concentrație optimă de apă în citoplasma corpului prin eliminarea excesului.

Comportamentul lui Paramecium Caudatum:
Răspunsurile lui Paramecium Caudatum la diferite tipuri de stimuli sunt învățate prin studierea reacțiilor sale și a grupării sau împrăștierii indivizilor într-o cultură. Răspunsul este pozitiv dacă animalul se deplasează spre un stimul și negativ când se îndepărtează. La un stimul negativ, animalul continuă să aibă o reacție de evitare până când scapă.
În reacția de evitare, bătaia ciliară se inversează, animalul se deplasează înapoi pe o distanță scurtă și apoi se rotește pe o traiectorie conică prin rotirea capătului anterior în direcția aborală în timp ce pivotează pe vârful posterior. Toate reglajele se fac prin încercare și eroare. Experimentele au arătat că extremitatea anterioară a animalului este mai sensibilă decât celelalte părți.
Răspunsurile lui Paramecium la diferiți stimuli pot fi grupate după cum urmează:
(i) Reacții la contact (Thigmotaxie):
Răspunsul la contact este variat la Paramecium. Dacă capătul anterior este atins ușor cu un vârf fin, apare o reacție puternică de evitare. Atunci când un Paramecium care înoată se ciocnește cu un obiect din apă, dar dacă este atins în altă parte este posibil să nu existe niciun răspuns. Un individ care se mișcă încet răspunde adesea pozitiv la contactul cu un obiect, oprindu-se pe acesta.
(ii) Reacții la substanțe chimice (chemotaxie):
În general, Paramecia răspunde la un stimul chimic prin evitarea reacției. Dacă se introduce o picătură de soluție slabă de sare (0,5 %) într-o populație de Paramecium pe o microlizieră, animalele răspund cu reacția de evitare și niciuna nu intră în picătură. Cu toate acestea, la acizi, răspunsul este pozitiv chiar și atunci când concentrația este suficient de puternică pentru a-i ucide.
(iii) Reacții la temperatură (termotaxie):
Paramecium caută o temperatură optimă cuprinsă între 24 și 28°C. Atunci când are loc o schimbare de temperatură cu mult peste sau sub intervalul optim, Paramecia prezintă o reacție de evitare. Căldura mai mare stimulează mișcările rapide și reacțiile de evitare până când animalele scapă sau sunt ucise.
(iv) Reacții la lumină (fototaxie):
Cu excepția Paramecium bursaria verde, care este pozitiv fototactic, celelalte specii sunt indiferente la lumina obișnuită. Cu toate acestea, atunci când intensitatea luminii este crescută brusc și brusc, urmează, în general, o reacție negativă. Paramecia prezintă un răspuns negativ imediat la razele ultraviolete.
(v) Reacții la curentul electric (galvanotaxie):
Paramecia răspunde la stimuli electrici. Atunci când doi electrozi sunt plasați unul în fața celuilalt într-o farfurie de mică adâncime care conține Paramecia și se aplică un curent constant, toate organismele înoată în aceeași direcție spre catodul sau electrodul negativ unde se concentrează în număr mare.
Dacă direcția curentului electric este inversată în timp ce Paramecia înoată spre catod, organismele inversează direcția și înoată spre noul catod.

(vi) Reacții la curentul de apă (Rheotaxis):
Paramecia prezintă o reotaxie pozitivă. Într-un curent de apă blând, Paramecia se va deplasa în cea mai mare parte odată cu curentul, cu capetele lor anterioare în amonte.
(vii) Reacții la gravitație (Geotaxie):
Paramecia prezintă, în general, un răspuns negativ la gravitație, după cum se observă într-o cultură în care mulți indivizi se adună aproape sub pelicula de suprafață, cu capetele lor anterioare în partea superioară. Dacă Paramecia este introdusă într-un tub în formă de U umplut cu apă și inversat, astupat la ambele capete, aceștia se deplasează imediat în sus în partea orizontală a tubului.
Reproducere la Paramecium Caudatam:
Paramecium Caudatum se reproduce asexuat prin fisiune binară transversală și suferă, de asemenea, mai multe tipuri de reorganizare nucleară, cum ar fi conjugarea, endomixia, autogamia, citogamia și hemixia etc.
(i) Fisiunea binară transversală:
Fisiunea binară transversală este cel mai frecvent tip de reproducere asexuată la Paramecium. Este un proces asexuat distinct și unic în care un specimen complet dezvoltat se împarte în doi indivizi fiică fără a lăsa un cadavru parental.
Planul de diviziune trece prin centrul celulei și se află într-un plan perpendicular pe axa lungă a corpului. Diviziunea corpului celular ca întreg este întotdeauna precedată de diviziunea nucleilor; într-adevăr, se pare că reproducerea este inițiată de activitatea și diviziunea nucleară.

Paramecium caudatum se reproduce prin fisiune binară transversală în condiții favorabile. În cazul fisiunii binare, micronucleul se împarte prin mitoză în două micronuclee fiice, care se deplasează la capetele opuse ale celulei. Macronucleul se alungește și se divide transversal prin amitoză.
Un alt citofaringe este dezmembrat și apar două noi vacuole contractile, unul lângă capătul anterior și altul lângă capătul posterior. Între timp, un șanț de constricție apare aproape de mijlocul corpului și se adâncește până când citoplasma este complet divizată.
Cele două paramecii „fiice” rezultate sunt de dimensiuni egale, fiecare conținând un set de organite celulare. Dintre cele două paramecii fiice produse, cea anterioară se numește proter și cea posterioară se numește opisthe. Acestea cresc până la dimensiunea maximă înainte de a avea loc o altă diviziune.
Procesul de fisiune binară necesită aproximativ două ore pentru a se finaliza și poate avea loc de una până la patru ori pe zi, producând între 2 și 16 indivizi. Într-un an se produc aproximativ 600 de generații.
Rata de înmulțire depinde de condițiile externe de hrană, temperatură, vârsta culturii și densitatea populației; de asemenea, de factorii interni de ereditate și fiziologie. În mod firesc, dacă toți descendenții unui individ ar supraviețui și s-ar reproduce, numărul de parameci produși ar fi în curând egal cu volumul pământului.
Termenul clonă este folosit pentru a se referi la toți indivizii care au fost produși de la un individ prin fisiune. Toți membrii unei clone sunt la fel din punct de vedere ereditar.
(ii) Conjugarea:
În mod obișnuit Paramecium Caudatum se înmulțește prin fisiune binară pentru perioade lungi de timp, dar la intervale de timp acest lucru poate fi întrerupt de unirea a două animale de-a lungul suprafețelor lor orale pentru procesul sexual de conjugare.
Conjugarea este definită ca fiind uniunea temporară a doi indivizi care fac schimb reciproc de material micronuclear. Este un tip unic de proces sexual în care două organisme se separă la scurt timp după schimbul de material nuclear.
Sonneborn (1947), pe baza comportamentului de împerechere al lui Paramecium Caudatum, a raportat că fiecare specie de Paramecium există într-un număr de varietăți sau syngens. Mai mult, în cadrul fiecărui syngen există un număr de tipuri de împerechere, de obicei două.
Tipurile de împerechere rămân identice din punct de vedere morfologic, dar prezintă diferențe fiziologice. La P. aurelia, există 14 syngens și 28 de tipuri de împerechere, în timp ce la P. caudatum există 16 syngens și 32 de tipuri de împerechere. S-a observat că, de obicei, paramecia nu se conjugă nici cu membrii propriului tip de împerechere, nici cu celelalte varietăți, ci doar cu cel de-al doilea tip de împerechere al propriei varietăți.

Factori care induc conjugarea:
Factorii care induc conjugarea variază de la o specie la alta, dar unii dintre ei sunt prezentați mai jos:
1. Conjugarea are loc, de obicei, în condiții de viață nefavorabile; se spune că înfometarea sau lipsa de hrană și o anumită dietă bacteriană sau anumite substanțe chimice induc procesul de conjugare la anumite specii de Paramecium.
2. Conjugarea are loc după aproximativ 300 de generații asexuate de fisiune binară, sau alternează cu fisiunea binară la intervale lungi de timp pentru a întineri clona muribundă, adică are loc la indivizii care trebuie să fi trecut printr-un număr dorit de generații asexuate, numită perioada de imaturitate, și apoi devin maturi din punct de vedere sexual pentru a se conjuga.
3. Conjugarea are loc atunci când are loc o schimbare în starea fiziologică a parameciilor, atunci are loc între astfel de indivizi care au dimensiuni ceva mai mici (210 microni lungime) și se află într-un stadiu care poate fi considerat ca o perioadă de bătrânețe nesănătoasă; paramecii de această condiție vor muri dacă nu li se permite să se conjuge.
4. Se spune că întunericul brusc în condiții de lumină și temperaturile scăzute induc procesul de conjugare la unele specii.
5. Conjugarea nu are loc în timpul nopții sau al întunericului; ea începe dimineața devreme și continuă până după-amiaza.
6. Se spune că o substanță proteică din cilia indivizilor de tip împerechere induce conjugarea.
Procesul de conjugare:
Procesul de conjugare diferă la diferite specii de Paramecium, dar relatarea de mai jos se referă la procesul de conjugare de la P. caudatum (Fig. 20.21).
În conjugare, două Paramecium caudatum (denumite pre-conjugante) de tipuri de împerechere opuse din aceeași varietate se apropie cu suprafețele lor ventrale și se unesc prin canelurile lor orale; cilia lor produc o substanță pe suprafața corpului care determină aderența celor două paramecii care se conjugă.
Se opresc din hrănire și aparatul lor de caneluri orale dispare. Pelicula și ectoplasma, în punctul de contact, ale ambelor se rup, iar între cele două animale se formează o punte protoplasmatică. Acum, acești indivizi se numesc conjuganți.
În această stare, perechea conjugantă înoată activ și simultan au loc o serie de modificări nucleare în fiecare conjugant, așa cum este descris mai jos:
Macronucleul începe să se dezintegreze, devine liber în textură și formează un schelet complex răsucit, în a doua jumătate a perioadei de conjugare el va dispărea în cele din urmă fiind absorbit în citoplasmă. Micronucleul fiecărui conjugant se divide de două ori, una dintre ele fiind o diviziune de reducere.
Astfel, în fiecare conjugant se produc patru micronuclee fiice haploide. Trei dintre acești patru micronuclee degenerează în fiecare, astfel încât rămâne doar unul singur.
Micronucleul rămas din fiecare conjugant se divide mitotic în doi pronuclei inegali sau nuclei gametici, formând un pro-nucleu feminin staționar mai mare și un pro-nucleu masculin migrator mai mic și mai activ.
Pro-nucleul migrator al unui conjugant traversează puntea protoplasmatică și fuzionează cu pro-nucleul staționar al celuilalt conjugant pentru a forma un nucleu sincariot sau nucleu de conjugare în care numărul diploid de cromozomi este restabilit și a avut loc un schimb de material ereditar.
Procesul a fost comparat cu fecundarea la animalele superioare, dar nu este vorba de fecundare deoarece nu sunt implicați gameți. Conjuganții se separă acum (după aproximativ 12-48 de ore) și se numesc ex-conjuganți. Sincariotul fiecărui ex-conjugant se divide de trei ori pentru a forma opt micronuclee în fiecare ex-conjugant.
Patru dintre cei opt micronuclee se măresc și devin macronuclee, iar trei dintre celelalte patru micronuclee dispar.
Micronucleul rămas se divide și, în același timp, ex-conjugantul se divide prin fisiune binară în două celule, fiecare având doi macronuclee și un micronuclee. Celulele și micronucleii lor se divid a doua oară pentru a forma patru parameci din fiecare ex-conjugant, astfel încât fiecare are un macronucleu și un micronucleu.
Noul macronucleu, ca și micronucleul, au fost făcute din material nou. Aceste noi nuclee conțin probabil un potențial nou și diferit, care se reflectă în indivizii sănătoși.
Semnificația conjugării:
O clonă se va stinge dacă nu are loc reorganizarea nucleară, dar clona poate fi întinerită pentru a-și recăpăta vigoarea de odinioară prin rearanjarea nucleară, această reorganizare nucleară este provocată de conjugare, astfel, conjugarea este esențială pentru continuarea fisiunii binare.
Semnificația conjugării a fost rezumată mai jos:
1. Conjugarea servește ca un proces de întinerire și reorganizare prin care se reface vitalitatea rasei. Dacă conjugarea nu are loc pentru perioade lungi de timp, paramecii slăbesc și mor. (Afirmația lui Woodruffs de a menține sănătoși paramecii timp de 22.000 de generații fără conjugare este infirmată de Sonneborn, deoarece a demonstrat că toți paramecii lui Woodruffs aparțineau aceluiași tip de împerechere).
2. Nu există nici o distincție de sex la conjuganți, deși doar paramecii din două tipuri de împerechere diferite din același soi se vor conjuga.
3. Nu există o distincție de sex, totuși pro-nucleul migrator activ este considerat ca fiind masculin, iar pro-nucleul staționar ca fiind feminin.
4. Conjugarea este doar o uniune temporară, nu există o fuziune a citoplasmei și nu se produce un zigot, dar nucleul fiecărui ex-conjugant conține material ereditar de la doi indivizi care se conjugă.
5. Conjugarea determină înlocuirea macronucleului cu material provenit de la sincariu, acesta este un eveniment de importanță fundamentală. În fisiunea binară, cromozomii macronucleului au fost distribuiți la întâmplare în celulele fiice, continuarea fisiunii binare a făcut ca clona să fie slabă, cu unele anomalii structurale.
Conjugarea duce la formarea numărului corect de cromozomi în macronucleu, astfel încât rasa se reînnoiește în vigoare. Rolul micronucleului este de a restabili un complex cromozomial și genetic echilibrat.
Comportamentul aberant în reproducere la Paramecium Caudatum:
Paramecium Caudatum prezintă anumite variații în comportamentul său nuclear în timpul fisiunii și conjugării, aceste deviații sunt endomixia, autogamia, citogamia și hemixia. În primele trei procese se realizează o recombinare genetică și se formează un nou macronucleu din micronucleu.
(i) Endomixia:
Woodruff și Erdmann (1914) au semnalat pentru prima dată un nou proces de reorganizare nucleară, endomixis (Gr., endon = înăuntru; mixis = amestecare) la Paramecium aurelia, o specie bimicronucleată (Fig. 20.22). Acest proces a fost descris ca având loc periodic, în care se produce un nou aparat macro-nuclear fără formarea de sincaryoni. Endomixisul are loc în cadrul unui singur individ.
Potrivit lui Woodruff și Erdmann, macronucleul degenerează și micronucleii se divid de două ori pentru a forma opt micronuclee. Șase micronuclee degenerează și rămân două. Rămânând doar doi micronuclee, animalul se divide prin fisiune în două celule, fiecare cu câte un micronucleu.
Micronucleul fiecărei celule fiice se divide de două ori pentru a forma patru micronuclee. Două micronuclee se măresc pentru a forma macronuclee. Animalul și micronucleii săi se divid astfel încât se formează doi indivizi fiică, fiecare având un macronucleu și doi micronuclee.
Endomixisul apare în acea varietate de P. aurelia care nu se conjugă, prin urmare, efectul endomixisului poate fi același cu cel al conjugării, deoarece ambele procese determină înlocuirea macronucleului cu material din micronucleu și ambele procese întineresc vitalitatea rasei.
Dar cele două procese diferă pentru că în endomixis nu există fuziune a pronucleilor; endomixisul poate fi comparat cu partenogeneza.
Cu toate acestea, unii lucrători au susținut, cu motive întemeiate, că endomixisul nu este un proces valabil și că a fost descris din cauza unor observații eronate. După toate probabilitățile, endomixisul nu are loc și poate fi doar un caz specializat de autogamie.

Mai târziu, Erdmamm și Woodruff (1916) au raportat endomixis la Paramecium caudatum. Cu toate acestea, Diller nu crede în validitatea acestui proces și consideră că Erdmann și Woodruff au combinat pur și simplu etapele de hemixie și autogamie într-o singură schemă, endomixis.
(ii) Autogamia:
Diller (1934, 1936) și Sonneborn (1950) au descris un proces de autofertilizare sau autogamie care are loc la un singur individ la Paramecium aurelia (Fig. 20.23). El a raportat că în autogamie trei diviziuni micro-nucleare (pregamice), care implică maturarea, produc nucleele gametice (pronucleii).
În timpul autogamiei la P. aurelia, cei doi micronuclee se divid de două ori (o dată meiotic) pentru a forma opt micronuclee, dintre care șase degenerează. Între timp, macronucleul crește într-o masă asemănătoare unui schelet care se rupe în bucăți pentru a fi ulterior absorbită în citoplasmă. Doi dintre cei opt micronuclee, sub formă de pronuclee, intră într-un con protoplasmatic bombat în apropierea gurii celulei.
Cei doi pronuclei fuzionează pentru a forma sincaryonul. Sincariotul se împarte de două ori pentru a forma patru micronuclee. Două micronuclee devin macronuclee. Parameciul și micronucleii săi se divid pentru a forma doi indivizi fiică, fiecare cu un macronucleu și două micronuclei. Acest proces se finalizează în aproximativ două zile.
Autogamia duce la întinerirea rasei.
Se aseamănă cu conjugarea în măsura în care noul macronucleu este format din material provenit din micronucleu, în noul macronucleu se restabilește numărul corect de cromozomi; și, de asemenea, prin faptul că are loc fuziunea a doi pronuclei. Dar autogamia se deosebește de conjugare pentru că la autogamie participă un singur individ care furnizează ambii pronuclei, este un fel de autofertilizare.
(iii) Hemixis:
Diller (1936) a semnalat hemixisul la Paramecium aurelia (Fig. 20.24). Hemixisul este în primul rând un proces de fragmentare și diviziune macro-nucleară fără nicio activitate micro-nucleară neobișnuită. Diller a clasificat hemixisul în patru tipuri, și anume A, B, C și D, așa cum se arată în Fig. 20.24 la P. aurelia, dar a întâlnit, de asemenea, toate tipurile în culturile de masă de P. caudatum și P. multimicronucleatum.
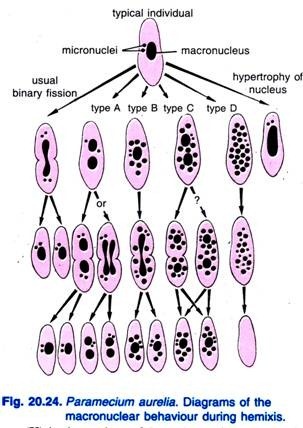
Tipul A este cea mai simplă formă de hemixie, caracterizată printr-o divizare a macronucleului în două sau mai multe părți. Această diviziune nu este sincronizată cu diviziunea micronucleară.
Tipul B se caracterizează prin extrudarea a una până la 20 sau mai multe bile de cromatină din macronucleu în citoplasmă.
Tipul C se caracterizează prin divizarea simultană a macronucleului în două sau mai multe porțiuni majore și prin extrudarea bilelor macro-nucleare în citoplasmă.
Se consideră că tipul D reprezintă condițiile patologice în care macronucleul suferă o fragmentare completă în bile de cromatină care în cele din urmă dispar din celulă. Micronucleii dispar în general înainte de dizolvarea macronucleului.
(iv) Citogamia:
Wichterman (1939) a raportat un alt proces sexual la Paramecium caudatum, pe care l-a numit citogamie. În cazul citogamiei, nu există un schimb nuclear. În acest proces, doi indivizi se apropie prin suprafețele lor ventrale, dar pelicula celor doi indivizi nu se desface.
Micronucleul fiecărui individ se divide de trei ori pentru a forma opt micronuclee, dintre care șase se dezintegrează în fiecare individ. Cele două micronuclee rămase fuzionează pentru a forma un sincariot în fiecare celulă. Animalele se separă acum.
Citogamia diferă de autogamie prin faptul că există două animale în contact unul cu celălalt, dar se aseamănă cu autogamia și conjugarea prin fuziunea a doi pronuclei. Citogamia se deosebește de conjugare prin faptul că nu există un schimb nuclear între cele două animale care se întâlnesc.
Câteva particule citoplasmatice raportate la Paramecium Caudatum:
(i) Particule Kappa:
În 1938, T.M. Sonneborn a raportat că unele rase (cunoscute ca ucigași sau tulpini ucigașe) de Paramecium produc o substanță otrăvitoare, numită paramecină, care este letală pentru alți indivizi numiți sensibili. Paramecina este solubilă în apă, difuzibilă și depinde pentru producerea ei de unele particule situate în citoplasma Parameciumului (tulpina ucigașă).
Aceste particule se numesc particule kappa. Particulele kappa au ADN și ARN. Un Paramecium ucigaș poate conține sute de particule kappa. Studiul detaliat al acestor particule a arătat că o genă dominantă (K) din nucleul Paramecium-ului este necesară pentru ca particulele kappa să existe, să se înmulțească și să producă paramecină.
(ii) mµ, Particule:
R.W. Siegel (1952) a raportat un alt tip de particule ucigașe în citoplasma unor Paramecium. Un Paramecium cu particule mp se numește ucigaș de pereche, deoarece atunci când se conjugă cu un Paramecium fără particule mµ, numit sensibil la pereche, atunci îl ucide pe acesta din urmă. Particulele mp sunt compuse, de asemenea, din ADN, ARN etc.
Aceste particule există numai la acei parameci al căror micronucleu conține cel puțin o genă dominantă a uneia dintre cele două perechi de gene cromozomiale nelegate (M1 și M2).
(iii) Particulele Pi:
Se presupune că aceste particule sunt forma mutantă a particulelor kappa, dar ele nu produc nici un fel de substanță toxică.
(iv) Particulele Lambda:
Aceste particule sunt semnalate la paramecii ucigași și se spune că produc o anumită substanță responsabilă pentru a provoca liza sau dezintegrarea paramecilor sensibili, adică care nu o posedă.