ADVERTISEMENTS:
In questo articolo parleremo di Paramecium Caudatum:- 1. Habitat, Habitat e Cultura di Paramecium Caudatum 2. Struttura di Paramecium Caudatum 3. Locomozione 4. Nutrizione 5. Respirazione ed escrezione 6. Osmoregolazione 7. Comportamento 8. Riproduzione 9. Comportamento aberrante nella riproduzione 10. Alcune particelle citoplasmatiche.
Contenuti:
- Habit, Habitat e Cultura del Paramecium Caudatum
- Struttura del Paramecium Caudatum
- Locomozione del Paramecium Caudatum
- Nutrizione nel Paramecium Caudatum
- Respirazione ed escrezione nel Paramecium Caudatum
- Osmoregolazione di Paramecium Caudatum
- Comportamento di Paramecium Caudatum
- Riproduzione in Paramecium Caudatum
- Comportamento aberrante nella riproduzione in Paramecium Caudatum
- Alcune particelle citoplasmatiche riportate in Paramecium Caudatum
- Habitat, Habitat e cultura del Paramecium Caudatum:
- Cultura di Paramecium:
- Struttura del Paramecium Caudatum:
- (i) Dimensioni e forma del Paramecium Caudatum:
- (ii) Pellicola:
- (iii) Cilia:
- (iv) Sistema Infraciliare:
- (v) Scanalatura orale e Citopigio:
- (vi) Citoplasma:
- (vii) Ectoplasma:
- (viii) Tricocisti:
- (ix) Neuromotorio e fibrille associate:
- (x) Endoplasma:
- (xi) Vacuoli contrattili:
- (xii) Vacuoli alimentari:
- Locomozione del Paramecium Caudatum:
- (i) Metabolia o contorsioni del corpo:
- (ii) Locomozione ciliare:
- Nutrizione del Paramecium Caudatum:
- Meccanismo di alimentazione:
- Digestione ed eduzione del Paramecium Caudatum:
- Respirazione ed escrezione del Paramecium Caudatum:
- Comportamento del Paramecium Caudatum:
- (i) Reazioni al contatto (tigmotassi):
- (ii) Reazioni alle sostanze chimiche (chemiotassi):
- (iii) Reazioni alla temperatura (Termotassi):
- (iv) Reazioni alla luce (fototassi):
- (v) Reazioni alla corrente elettrica (galvanotassi):
- (vi) Reazioni alla corrente d’acqua (Reotassi):
- (vii) Reazioni alla gravità (Geotassi):
- Riproduzione in Paramecium Caudatam:
- (i) Fissione binaria trasversale:
- (ii) Coniugazione:
- Comportamento aberrante nella riproduzione nel Paramecium Caudatum:
- (i) Endomissia:
- (ii) Autogamia:
- (iii) Emixis:
- (iv) Citogamia:
- Alcune particelle citoplasmatiche riportate nel Paramecium Caudatum:
- (i) Particelle Kappa:
- (ii) mµ, particelle:
- (iii) Particelle Pi:
- (iv) Particelle Lambda:
Habitat, Habitat e cultura del Paramecium Caudatum:
Paramecium caudatum (Gr., paramekes = oblungo; L., caudata = coda) si trova comunemente in stagni d’acqua dolce, piscine, fossati, ruscelli, laghi, serbatoi e fiumi. Si trova specialmente in abbondanza negli stagni stagnanti ricchi di materia in decomposizione, negli infusi organici e nelle acque di scarico. Il Paramecium caudatum è un organismo a vita libera e questa specie ha una distribuzione mondiale.
Cultura di Paramecium:
ADVERTISEMENTS:
Prendere delle erbacce sommerse da uno stagno e metterle in un vaso di acqua distillata, coprire il vaso e lasciarlo marcire; in pochi giorni appariranno degli sciami di Paramecia. Ora fate bollire il fieno nell’acqua, decantate l’infuso e aggiungete qualche chicco di grano, e lasciatelo riposare fino a quando sarà torbido di batteri.
Trasferire le Paramecia dal primo vaso in questo liquido dove si moltiplicheranno rapidamente. Le infusioni di fieno da sole produrranno Paramecia mostrando la presenza di cisti, e sono state riportate cisti simili a granelli di sabbia, ma non c’è alcuna prova che i Paramecia formino cisti, poiché non sono mai state confermate.
Struttura del Paramecium Caudatum:
(i) Dimensioni e forma del Paramecium Caudatum:
Paramecium caudatum (Fig. 20.1) è un organismo microscopico e visibile ad occhi nudi come un corpo minuto e allungato.
ADVERTISIMENTI:
Appare grigio chiaro o bianco e misura comunemente tra 170 e 290 micron di lunghezza e può raggiungere una lunghezza fino a 300-350 micron. P. caudatum assomiglia alla suola di una pantofola o di una scarpa, quindi l’animale è comunemente conosciuto come animaletto da pantofola. È quattro volte più lungo che largo e un po’ cilindrico con estremità nettamente diverse.
La parte anteriore che si muove in avanti è snella con un’estremità smussata o arrotondata, mentre l’estremità posteriore è un po’ appuntita o a forma di cono. La parte più larga dell’organismo è appena sotto la metà. Il corpo dell’animale è di forma asimmetrica e presenta una superficie orale o ventrale ben definita e una aborale o dorsale.
(ii) Pellicola:
Il corpo è coperto da una sottile pellicina di gelatina a doppio strato, elastica e soda. La pellicina mantiene la forma dell’animale ma è abbastanza elastica da permettere le contrazioni. La pellicola ha una doppia membrana, la membrana esterna è continua con le ciglia e la membrana interna con l’ectoplasma. Sotto l’ingrandimento maggiore del microscopio, la pellicina mostra depressioni rettangolari o esagonali sulla sua superficie.

Questa disposizione si conserva sulla superficie dorsale del Paramecium ma sulla superficie ventrale le creste convergono davanti e dietro verso un’apertura preorale e postorale. Ogni depressione esagonale è perforata da un’apertura centrale attraverso la quale emerge un singolo cilio. I margini anteriori e posteriori delle depressioni esagonali portano le aperture delle tricocisti.
Lo studio al microscopio elettronico della pellicola (Fig. 20.2) di Ehret e Powers (1957) ha rivelato che le depressioni esagonali corrispondono a serie regolari di cavità, gli alveoli. Tutti gli alveoli formano collettivamente uno strato alveolare continuo, che è delimitato da una membrana alveolare esterna e una interna.
Lo strato esterno si trova in stretto contatto sotto la membrana cellulare esterna. Pertanto, la pellicina comprende la membrana cellulare esterna, la membrana alveolare esterna e la membrana alveolare interna.
(iii) Cilia:
L’intero corpo è coperto da numerose, piccole proiezioni simili a capelli chiamate ciglia. Le ciglia si presentano in file longitudinali su tutto il corpo, questa condizione è conosciuta come olotria in cui le ciglia del corpo sono uguali. Le cilia hanno la stessa struttura dei flagelli, hanno una guaina protoplasmatica esterna o membrana plasmatica con nove doppie fibrille longitudinali in un anello periferico. In alcune cilia le nove fibrille esterne non sono appaiate.
ADVERTIME:
Ci sono due fibrille longitudinali centrali che sono più sottili delle fibrille esterne. Ogni cilium nasce da un granulo basale o cinetosoma. Le nove coppie di fibrille periferiche si fondono insieme per formare la parete del cinetosoma, quindi, il cinetosoma è un tubo che è aperto o chiuso alla sua estremità inferiore, le due fibrille centrali si fermano a livello della pellicina nella maggior parte dei ciliati.
Dal cinetosoma nasce un sottile rizoplasto che non si unisce al nucleo. Anche molti Metazoi hanno cilia, la loro struttura è la stessa, tranne che il granulo basale è diverso e ha sottili filamenti o fibre radicali che si estendono nel citoplasma. Ma le ciglia differiscono dai flagelli per essere generalmente più numerose e di dimensioni inferiori.
La ciliatura può essere convenientemente divisa in cilia corporee o somatiche che si trovano sulla superficie del corpo, e in cilia orali che sono associate alla regione della bocca. Le ciglia del corpo sono uguali, ma sono più lunghe all’estremità posteriore, da cui il nome caudatum. Le ciglia sono organelli di locomozione e di raccolta del cibo, agiscono anche come recettori sensoriali e rilevano gli stimoli dell’ambiente esterno.

Ultra struttura delle ciglia:
Le ciglia e i flagelli hanno una composizione fibrillare. Alla base il cilio ha il diametro di circa 0,2 micron o 2.000 A0 che può essere fino a 10 micron sopra la superficie cellulare. Le ciglia sono delimitate da una membrana unitaria di 90 A0 di spessore che assomiglia e rimane continua con la membrana plasmatica. Lo spazio delimitato del cilio contiene una sostanza acquosa nota come matrice.
Nella matrice, rimangono incorporate undici fibrille longitudinali o microtubuli. Delle undici fibrille, due si trovano al centro, mentre le restanti nove fibrille rimangono disposte perifericamente intorno alle fibrille centrali. Ciascuna delle nove fibrille esterne ha un diametro di 360A0 ed è composta da due sub-fibrille di diametro da 180 a 250A0.
Queste sub-fibrille sono designate come sub-fibrilla A e sub-fibrilla B.
ADVERTISIMENTI:
La subfibrilla A è leggermente più grande della subfibrilla B. La subfibrilla A emette due spesse proiezioni o braccia da un lato. Le braccia della sottofibrilla A di tutte le fibrille esterne rimangono dirette in senso orario. Inoltre, la sub-fibrilla A è più vicina al centro del cilium rispetto alla sub-fibrilla B. Entrambe le sub-fibrille hanno una parete comune di 50A0 di spessore.
Le due fibrille centrali non hanno sub-fibrille appaiate come le nove fibrille periferiche, ma ciascuna contiene un solo tubulo. Ogni fibrilla centrale ha un diametro di circa 250A0 ed è composta da una parete spessa 60A0.
Entrambe le fibrille centrali rimangono separate da uno spazio di 350A0 e rimangono avvolte da una guaina comune. Gibbnos (1967) ha riportato che la guaina delle fibrille centrali fornisce nove collegamenti orientati radialmente o raggi a ciascuna sub-fibrilla A.

La microscopia elettronica ad alta risoluzione ha rivelato che ciascuna delle fibrille periferiche e centrali delle ciglia e dei flagelli è composta da dieci a dodici filamenti di 40A0 di spessore. Ogni filamento è a perline. Ogni perlina rimane disposta in reticoli di 40 per 50A0 nel piano della parete del tubulo. Queste perline sono considerate come la subunità di base della struttura del tubulo.
(iv) Sistema Infraciliare:
Il sistema infraciliare si trova appena sotto gli alveoli pellicolari. È costituito dal cinetosoma o corpo basale e dal cinetodesma. Le ciglia nascono dai cinetosomi e da ogni cinetosoma nasce una delicata fibrilla citoplasmatica chiamata cinetodesma, (Fig. 20.2).
Sotto la pellicina leggermente a destra, ma unita a tutti i cinetosomi di una fila longitudinale, si trova un fascio longitudinale di diversi cinetosomi, un cinetosoma di ogni cinetosoma si estende per una distanza anteriore nel proprio fascio di cinetosomi.
Una fila longitudinale di cinetosomi con i loro cinetosomi forma un’unità longitudinale chiamata cinetia. Tutte le cinetie o cinetie formano il sistema infraciliare di un ciliato. I cinetismi si trovano nella corteccia sotto la pellicina, il loro numero è abbastanza costante per ogni ciliato.
Il sistema infra-ciliare controlla e coordina i movimenti delle ciglia, e porta alla formazione di organelli nella divisione cellulare, ad esempio, alcuni cinetidi formano la bocca. Nella fissione binaria dei ciliati i cinetidi sono tagliati trasversalmente in due, ognuno dei quali va ad una cellula figlia, questa è chiamata fissione pericinetale.
(v) Scanalatura orale e Citopigio:
Sul lato ventrolaterale c’è una grande depressione obliqua e poco profonda chiamata solco orale o peristoma che dà all’animale un aspetto asimmetrico. Corre obliquamente all’indietro da un lato (di solito da sinistra a destra ma in alcuni casi da destra a sinistra) e termina un po’ dietro il corpo centrale. Il solco orale sfocia in una breve depressione conica a forma di imbuto chiamata vestibolo.
ADVERTIME:
Il vestibolo conduce direttamente all’apertura fissa di forma ovale chiamata citostoma (bocca). Dal citostoma si estende direttamente verso il centro del corpo l’ampio citofaringe. Il citofaringe poi gira bruscamente verso il lato posteriore per diventare l’esofago sottile e affusolato.
Quindi, l’esofago è approssimativamente parallelo alla superficie del corpo di Paramecium, tranne che alla sua estremità posteriore. Qui l’esofago gira di nuovo verso il centro dell’animale per condurre nel vacuolo alimentare in formazione.
Il citopigio (chiamato anche ano cellulare o macchia anale o citopratto) si trova sulla superficie ventrale del corpo quasi verticalmente dietro il citostoma o bocca. Le particelle di cibo non digerite vengono eliminate attraverso il citopigio. La conciliazione del citofaringe è molto complicata.
Gelei (1934) ha riportato la presenza di quattro file e Lund (1941) ha osservato almeno quattro file. Una struttura chiamata penniculus si trova sulla parete sinistra del citofaringe e gira a spirale per circa 90 gradi in modo che la sua estremità posteriore sia sulla superficie orale (ventrale) dell’esofago.
Secondo Lund, il pennicolo è composto da otto file di cilia disposte in due blocchi strettamente impostati di ciascuna. Una banda simile composta da quattro file di cilia lunghe e meno compatte che nel penniculus è chiamata quadrulus. Si estende a spirale lungo la parete dorsale della cavità buccale e termina vicino al pennicolo.
Il penniculus e il quadrulus sono stati erroneamente chiamati una membrana ondulata da alcuni lavoratori. Il quadrulus e il penniculus controllano il passaggio del cibo. Non si sa come funzionano le ciglia, probabilmente le loro fibrille si contraggono in modo ritmico che provoca la flessione. Gelei (1925) ha sottolineato che la funzione del penniculus è la forzatura degli elementi alimentari nel corpo.

(vi) Citoplasma:
Il citoplasma è differenziato in una zona stretta, esterna o corticale chiamata ectoplasma e una regione più grande, interna o midollare chiamata endoplasma.
(vii) Ectoplasma:
L’ectoplasma (ectosarco o corteccia) è una parte permanente del corpo, nettamente delimitata dall’endoplasma. L’ectoplasma forma uno strato esterno solido, chiaro, sottile e denso. Contiene le tricocisti, le ciglia e le strutture fibrillari ed è delimitato esternamente da un rivestimento chiamato pellicina.
(viii) Tricocisti:
Inserite nell’ectoplasma ad angolo retto rispetto alla superficie ci sono piccole sacche a forma di fuso chiamate tricocisti. Un piccolo punto su ogni margine anteriore e posteriore dell’esagono segna la posizione di una tricocisti. Sono pieni di un fluido denso e rifrangente che ha una sostanza rigonfiante, all’estremità esterna c’è una testa conica o spiga.
Le tricocisti giacciono perpendicolari nell’ectoplasma, si aprono tramite piccoli pori sulle creste delle aree esagonali della pellicola.
Sorgono dai cinetosomi delle ciglia, poi migrano e si localizzano a uguale distanza nell’endoplasma. Quando l’animale è irritato, le tricocisti vengono scaricate come lunghi fili appiccicosi. Una tricocisti scaricata ha una punta opaca, simile a un’unghia rovesciata, e un lungo fusto striato, ma il fusto non si vede nello stato non scaricato e si forma probabilmente durante la scarica.
La funzione dei tricocisti è incerta, ma vengono scaricati come reazione a contatti locali e lesioni, potrebbero servire come organelli di difesa.
Ma questo è incerto perché le tricocisti sono inefficaci contro il Didinium, il principale predatore del Paramecium, potrebbero servire per fissare l’animale ad un punto durante l’alimentazione. In alcuni ciliati, le tricocisti agiscono come organelli di offesa. Dopo lo scarico delle tricocisti, rigenerate dai cinetosomi.


(ix) Neuromotorio e fibrille associate:
Secondo Lund (1933) sulla parete dorsale sinistra del citofaringe a circa il livello del margine posteriore del citostoma si trova una massa molto piccola e bilobata, il neuromotorio. Dal neuromotorio, le fibrille si irradiano nell’endoplasma.
Di queste quattro o più di solito passano quasi alla parete dorsale del corpo, ma le altre sono più corte e non definite nella posizione. Tutte sono chiamate fibrille endoplasmatiche. La loro funzione è sconosciuta, ma possono coordinare i movimenti di alimentazione delle ciglia orali. Le fibrille possono anche dare supporto meccanico, elasticità, contrattilità, conduttività e influenza metabolica.

(x) Endoplasma:
L’endoplasma o midollo è la parte più fluida e voluminosa del citoplasma che contiene molti granuli citoplasmatici e altre inclusioni e strutture di natura specializzata. Le inclusioni citoplasmatiche sono mitocondri, apparati di Golgi, vacuoli, cristalli, granuli e cromidi, ecc. Altre strutture, cioè nuclei, vacuoli contrattili e vacuoli alimentari si trovano anche nell’endoplasma.
Nuclei:
Nell’endoplasma vicino al citostoma si trovano due nuclei, cioè il Paramecium è eterocariota, un grande macronucleo ellissoidale e granulare e un altro piccolo micronucleo compatto.
Il macronucleo è un corpo cospicuo, ellissoidale o reniforme. È di tipo compatto e contiene fili sottili e granuli di cromatina discreti e strettamente impacchettati di dimensioni variabili, incorporati in una matrice acromatica. Possiede molti nucleoli e molto più materiale cromatinico (DNA).
È un nucleo somatico o vegetativo. Si divide amitoticamente e controlla le funzioni vegetative (attività metaboliche) dell’animale. Subisce la mitosi.
Il micronucleo è piccolo, compatto e sferico. Si trova generalmente vicino al macronucleo, spesso in una concavità. Granuli di cromatina fine e fili sono distribuiti uniformemente in tutta la struttura. Il micronucleo si divide mitoticamente e controlla la riproduzione.
Il numero di micronuclei varia con la specie; è uno in P. caudatum, due in P. aurelia e molti in P. multimicronucleatum. Il micronucleo contiene un nucleo distinto in P. aurelia ma non si trova in P. caudatum. Moses (1949; 1950) ha riportato che il macronucleo e il micronucleo sono identici nella composizione chimica.
(xi) Vacuoli contrattili:
Ci sono due grandi vacuoli contrattili pieni di liquido, ciascuno situato vicino a un’estremità del corpo vicino alla superficie dorsale. La loro posizione è fissa (a differenza dell’Amoeba), si trovano tra l’ectoplasma e l’endoplasma, ma sono organelli temporanei che scompaiono periodicamente. In alcune specie, sembrano avere una membrana di rivestimento, nel qual caso non scompaiono completamente durante la sistole.
Collegati ad ogni vacuolo contrattile ci sono da cinque a dodici canali tubolari a raggiera, ciascuno composto da una parte terminale, una lunga ampolla che collassa quando è vuota, e un breve canale iniettore che si apre nel vacuolo.

I canali comunicano con gran parte del corpo da dove prendono liquidi e li versano nel vacuolo che così si ricostituisce e cresce di dimensioni, quando il vacuolo contrattile raggiunge la sua dimensione massima si contrae improvvisamente (sistole) e scarica il suo contenuto attraverso un poro permanente nella pellicola, poi i canali formano nuovamente i vacuoli contrattili, i canali non scompaiono del tutto poiché sono strutture permanenti.
I due vacuoli contrattili scaricano irregolarmente, quello posteriore si contrae più rapidamente perché è vicino alla citofaringe e vi entra più acqua. La funzione principale dei canali e dei vacuoli contrattili è idrostatica, rimuovono l’eccesso di acqua dal protoplasma, l’acqua viene in parte assorbita e in parte assunta durante l’alimentazione.
La sostanza di scarto azotata ha composti di ammoniaca e alcuni urati che vengono espulsi dai vacuoli contrattili insieme a CO2, ma non c’è alcuna prova che la materia escretrice sia secreta dal protoplasma nei canali o nei vacuoli contrattili.
C’è una citofisi fissa permanente o un ano sotto il peristoma a un lato del citofaringe, attraverso il quale vengono espulsi resti di cibo non digeriti e urati.
Lo studio al microscopio elettronico dei vacuoli contrattili ha rivelato che ogni apparato contrattile è costituito da alcuni tubuli del reticolo endoplasmatico, tubuli nefridiali, canali di alimentazione, vacuoli accessori (canali radiali) e vacuolo contrattile principale. Si suppone che i vacuoli accessori siano le ampolle dei canali di alimentazione (Fig. 20.11).
(xii) Vacuoli alimentari:
Sono corpi non contrattili, approssimativamente sferici, di dimensioni e numero variabili, che giacciono nell’endoplasma. Contengono particelle di cibo ingerito, principalmente batteri e una piccola quantità di fluido delimitato da una sottile membrana definita. Volkonsky (1934) ha proposto il nome di gastrioli per questi vacuoli. Associati ai vacuoli alimentari sono i granuli digestivi.
Locomozione del Paramecium Caudatum:
Paramecium Caudatum esegue la locomozione con due metodi, cioè il metabolismo o le contorsioni del corpo e le ciglia.
(i) Metabolia o contorsioni del corpo:
Il corpo del Paramecium Caudatum possiede elasticità, può schiacciarsi attraverso un passaggio più stretto del suo corpo, dopo di che il corpo assume la sua forma normale. Questo cambiamento temporaneo della forma del corpo è la metabolizzazione, che viene effettuata nel Paramecium dal protoplasma.
(ii) Locomozione ciliare:
La locomozione portata dalle ciglia è il metodo principale. Le ciglia possono battere in avanti o all’indietro permettendo all’animale di nuotare anteriormente o posteriormente.
Normalmente l’animale nuota in avanti, le ciglia battono all’indietro ma obliquamente, le ciglia si irrigidiscono e si piegano rapidamente all’indietro fino quasi a toccare la superficie del corpo, questa è chiamata la corsa effettiva; poi le ciglia si afflosciano e ritornano lentamente alla posizione verticale originale, questa è chiamata la corsa di recupero.
Le ciglia della stessa fila trasversale battono insieme e quelle della stessa fila longitudinale battono una dopo l’altra dall’estremità anteriore a quella posteriore.

Questo movimento coordinato di cilia è chiamato ritmo metacronale, che è dovuto al sistema infracciliare; questo provoca il nuoto in avanti dell’animale. Ma quando le ciglia del corpo battono obliquamente all’indietro, allora allo stesso tempo le ciglia più lunghe del solco orale battono più vigorosamente, il che provoca la deviazione a sinistra dell’estremità anteriore.
L’azione delle ciglia del corpo e del solco orale fa ruotare l’animale sul suo asse lungo. Questa rotazione è sempre verso sinistra (tranne in P. calkinsi che ruota a spirale verso destra).
Questa combinazione di movimento in avanti, sterzata e rotazione fa muovere l’animale in avanti in un percorso a spirale in senso antiorario. Questo percorso ha un asse dritto, e la stessa superficie corporea dell’animale rimane verso l’asse del percorso a spirale. Ma nel nuotare all’indietro, tutte le specie ruotano verso destra.
Il battito ciliare può essere invertito in modo che le ciglia si muovano obliquamente in avanti con cui l’animale nuota all’indietro. Con l’azione ciliare, il Paramecium si muove con una velocità di 1500 micron o più al secondo.
Jennings ha sostenuto che la spiralizzazione del Paramecium è dovuta al fatto che mentre le ciglia colpiscono principalmente all’indietro, lo fanno obliquamente a destra causando così il rotolamento dell’animale a sinistra.
Anche questa deviazione del corpo verso la superficie aborale è dovuta in gran parte alla maggiore potenza del colpo effettivo delle ciglia orali che colpiscono più direttamente all’indietro. Il risultato – la rotazione del Paramecio sul suo asse lungo – permette al Paramecio di seguire un percorso più o meno rettilineo nel formare grandi spirali.
Nutrizione del Paramecium Caudatum:
Nel Paramecium Caudatum, la nutrizione è olozoica. Il cibo comprende principalmente batteri e piccoli protozoi. Il Paramecium non aspetta il cibo ma lo caccia attivamente.
Si sostiene che il Paramecium Caudatum mostri una scelta nella selezione del suo cibo, ma non sembrano esserci basi per questo, anche se inghiotte solo alcuni tipi di batteri; i dati disponibili suggeriscono che da 2 a 5 milioni di individui di Bacillus coli sono divorati da un singolo Paramecium in 24 ore. Si nutre anche di piante unicellulari come alghe, diatomee, ecc. e piccoli pezzi di animali e vegetali.
Meccanismo di alimentazione:
Quando il Paramecium Caudatum entra in una regione di cibo abbondante, si riposa. Si nutre solo a riposo o quando nuota molto lentamente, non si nutre mai quando nuota velocemente. Il battito delle ciglia del solco orale fa sì che un vortice a forma di cono di acqua carica di cibo venga spazzato nel solco orale da una distanza anteriore all’estremità anteriore (Fig. 20.16).
Le particelle di cibo passano poi al vestibolo da dove alcune particelle di cibo vengono respinte e gettate fuori, ma altre passano nel citostoma.
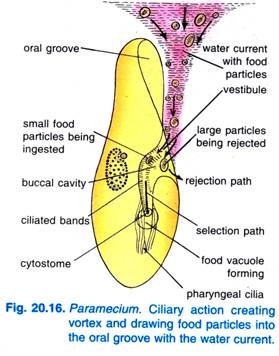
Al termine del citofaringe, si forma un vacuolo alimentare che si riempie di particelle di cibo. Il quadrulo e i peniculi controllano il passaggio del cibo nel vacuolo alimentare che si forma lateralmente. Quando il vacuolo alimentare raggiunge una certa dimensione, le fibre post-buccali afferrano il vacuolo alimentare e questo viene pizzicato da esse e riprende il suo corso.
Il vacuolo contiene dell’acqua oltre al cibo. Movimenti rotatori dell’endoplasma chiamati ciclosi portano i vacuoli alimentari lungo un percorso definito che è funzionalmente equivalente a un tratto digestivo.
Il tratto inizia dalla fine della citofaringe, poi verso la parte posteriore, poi in avanti per circolare con l’endoplasma, poi verso la superficie dorsale, poi verso l’estremità anteriore, poi verso il basso fino al citopigio. All’inizio del suo viaggio il vacuolo alimentare diminuisce di dimensioni, poi aumenta di nuovo.

Digestione ed eduzione del Paramecium Caudatum:
Durante la ciclosi, la digestione avviene tramite enzimi secreti dal protoplasma nei vacuoli. Nella digestione, le proteine vengono trasformate in aminoacidi, i carboidrati in zuccheri solubili e glicogeno, e probabilmente anche i grassi vengono digeriti.
Il contenuto dei vacuoli alimentari è inizialmente acido (pH circa 4) e poi diventa alcalino, la digestione principale avviene durante la fase alcalina. La materia non digerita viene espulsa attraverso il citopigio con una certa forza.
La ciclosi può essere dimostrata sperimentalmente; se il latte colorato con rosso Congo viene somministrato al Paramecium, i globuli di grasso del latte nei vacuoli alimentari diventeranno prima rossi a causa della reazione acida degli enzimi, poi passeranno dalle tonalità del viola al blu a causa della reazione alcalina, i vacuoli mostreranno il corso della ciclosi.
Respirazione ed escrezione del Paramecium Caudatum:
Lo scambio di gas (ossigeno e anidride carbonica) avviene attraverso la pellicola semipermeabile come altri protozoi d’acqua dolce attraverso il processo di diffusione. Il Paramecium Caudatum ottiene il suo ossigeno dall’acqua circostante. L’anidride carbonica e i rifiuti organici come l’ammoniaca derivanti dal metabolismo sono probabilmente espulsi diffondendo verso l’esterno nell’acqua in direzione opposta.

6. Osmoregolazione in Paramecium Caudatum:
Paramecium Caudatum ha due vacuoli contrattili, uno anteriore e uno posteriore. La funzione dei vacuoli contrattili è l’osmoregolazione, cioè, regolare il contenuto di acqua del corpo e può servire anche nell’escrezione di rifiuti azotati come l’urea e l’ammoniaca.
L’eccesso di acqua (a causa della continua endosmosi) all’interno del citoplasma viene secreto nei tubuli del reticolo endoplasmatico e va nei tubuli nefridiali → canali di alimentazione → e si raccoglie nell’ampolla di una serie di 6-11 canali a raggiera che convergono verso e scaricano in ogni vacuolo. I canali sono più cospicui quando un vacuolo si sta formando.
Quando ogni vacuolo si gonfia (diastole) fino a una certa dimensione, si contrae (sistole) e scarica all’esterno probabilmente attraverso un poro. I vacuoli contrattili si contraggono alternativamente, a intervalli di 10 – 20 secondi.
Il vacuolo contrattile posteriore lavora più velocemente del vacuolo anteriore a causa dell’assunzione di una grande quantità di acqua nella regione posteriore da parte della citofaringe. I vacuoli contrattili mantengono una concentrazione ottimale di acqua nel citoplasma del corpo smaltendo l’eccesso.

Comportamento del Paramecium Caudatum:
Le risposte del Paramecium Caudatum a vari tipi di stimoli vengono apprese dallo studio delle sue reazioni e del raggruppamento o dispersione degli individui in una cultura. La risposta è positiva se l’animale si muove verso uno stimolo e negativa quando si allontana. Ad uno stimolo negativo l’animale continua a dare la reazione di evitamento fino a quando non fugge.
Nella reazione di evitamento, il battito ciliare si inverte, l’animale si muove all’indietro per una breve distanza, e poi ruota in un percorso conico facendo oscillare l’estremità anteriore aboralmente mentre ruota sulla punta posteriore. Tutte le regolazioni sono fatte per tentativi ed errori. Gli esperimenti hanno dimostrato che l’estremità anteriore dell’animale è più sensibile delle altre parti.
Le risposte del Paramecium a diversi stimoli possono essere raggruppate come segue:
(i) Reazioni al contatto (tigmotassi):
La risposta al contatto è varia nel Paramecio. Se l’estremità anteriore viene leggermente toccata con una punta sottile, si verifica una forte reazione di evitamento. Quando un Paramecium che nuota si scontra con qualche oggetto nell’acqua, ma se viene toccato altrove può non esserci alcuna risposta. Un individuo che si muove lentamente spesso risponde positivamente al contatto con un oggetto venendo a posarsi su di esso.
(ii) Reazioni alle sostanze chimiche (chemiotassi):
Generalmente le Paramecia rispondono ad uno stimolo chimico evitando la reazione. Se una goccia di soluzione salina debole (0,5 per cento) viene introdotta in una popolazione di Paramecium su un micro-slide, gli animali rispondono con la reazione di evitamento e nessuno entra nella goccia. Agli acidi, tuttavia, la risposta è positiva anche quando la concentrazione è di forza sufficiente per ucciderli.
(iii) Reazioni alla temperatura (Termotassi):
Paramecium cerca una temperatura ottimale da 24 a 28°C. Quando si verifica un cambiamento di temperatura marcatamente al di sopra o al di sotto dell’intervallo ottimale, i parameci mostrano una reazione di evitamento. Un calore maggiore stimola il movimento rapido e le reazioni di evitamento finché gli animali non fuggono o vengono uccisi.
(iv) Reazioni alla luce (fototassi):
Con l’eccezione del Paramecium bursaria verde, che è positivamente fototattico, le altre specie sono indifferenti alla luce ordinaria. Tuttavia, quando l’intensità della luce è improvvisamente e bruscamente aumentata, segue generalmente una reazione negativa. I parameci mostrano una risposta negativa immediata ai raggi ultravioletti.
(v) Reazioni alla corrente elettrica (galvanotassi):
Paramecia risponde agli stimoli elettrici. Quando due elettrodi sono posti uno di fronte all’altro in un piatto poco profondo contenente Paramecia e viene applicata una corrente costante, tutti gli organismi nuotano nella stessa direzione verso il catodo o elettrodo negativo dove si concentrano in gran numero.
Se la direzione della corrente elettrica viene invertita mentre le Paramecia nuotano verso il catodo, gli organismi invertono la direzione e nuotano verso il nuovo catodo.

(vi) Reazioni alla corrente d’acqua (Reotassi):
I parameci mostrano una reotassi positiva. In una leggera corrente d’acqua le Paramecia si muoveranno per lo più con il flusso, con le loro estremità anteriori a monte.
(vii) Reazioni alla gravità (Geotassi):
Le Paramecia generalmente mostrano una risposta negativa alla gravità, come si vede in una cultura in cui molti individui si riuniscono sotto la pellicola superficiale con le loro estremità anteriori in alto. Se le Paramecia vengono introdotte in un tubo a forma di U rovesciato pieno d’acqua e tappato alle due estremità, si muovono immediatamente verso l’alto nella parte orizzontale del tubo.
Riproduzione in Paramecium Caudatam:
Paramecium Caudatum si riproduce asessualmente per fissione binaria trasversale e subisce anche diversi tipi di riorganizzazione nucleare, come coniugazione, endomissia, autogamia, citogamia ed emixia, ecc.
(i) Fissione binaria trasversale:
La fissione binaria trasversale è il tipo più comune di riproduzione asessuata nel Paramecium. Si tratta di un processo asessuale distintamente unico in cui un esemplare completamente cresciuto si divide in due individui figli senza lasciare un cadavere dei genitori.
Il piano di divisione è attraverso il centro della cellula e in un piano ad angolo retto rispetto all’asse lungo del corpo. La divisione del corpo cellulare nel suo insieme è sempre preceduta dalla divisione dei nuclei; sembra infatti che la riproduzione sia iniziata dall’attività e dalla divisione nucleare.

Paramecium Caudatum si riproduce per fissione binaria trasversale in condizioni favorevoli. Nella fissione binaria, il micronucleo si divide per mitosi in due micronuclei figli, che si spostano alle estremità opposte della cellula. Il macronucleo si allunga e si divide trasversalmente per amitosi.
Un altro citofaringe si stacca e appaiono due nuovi vacuoli contrattili, uno vicino all’estremità anteriore e un altro vicino a quella posteriore. Nel frattempo, un solco di costrizione appare vicino al centro del corpo e si approfondisce fino a quando il citoplasma è completamente diviso.
I due parameci “figli” risultanti sono di dimensioni uguali, ciascuno contenente un insieme di organelli cellulari. Delle due parameci figlie prodotte, quella anteriore è chiamata proter e quella posteriore è chiamata opisto. Crescono fino alla dimensione massima prima che avvenga un’altra divisione.
Il processo di fissione binaria richiede circa due ore per essere completato e può avvenire da una a quattro volte al giorno, producendo da 2 a 16 individui. Circa 600 generazioni sono prodotte in un anno.
Il tasso di moltiplicazione dipende dalle condizioni esterne di cibo, temperatura, età della cultura e densità della popolazione; anche dai fattori interni di ereditarietà e fisiologia. Naturalmente, se tutti i discendenti di un individuo dovessero sopravvivere e riprodursi, il numero di paramecia prodotti sarebbe presto pari al volume della terra.
Il termine clone è usato per indicare tutti gli individui che sono stati prodotti da un individuo per fissione. Tutti i membri di un clone sono ereditari allo stesso modo.
(ii) Coniugazione:
Ordinariamente Paramecium Caudatum si moltiplica per fissione binaria per lunghi periodi di tempo, ma a intervalli questo può essere interrotto dall’unione di due animali lungo le loro superfici orali per il processo sessuale di coniugazione.
La coniugazione è definita come l’unione temporanea di due individui che si scambiano reciprocamente materiale micro nucleare. È un tipo unico di processo sessuale in cui due organismi si separano subito dopo lo scambio di materiale nucleare.
Sonneborn (1947), sulla base del comportamento di accoppiamento di Paramecium Caudatum, ha riportato che ogni specie di Paramecium esiste in un certo numero di varietà o singen. Inoltre, all’interno di ogni syngen ci sono un certo numero di tipi di accoppiamento, di solito due.
I tipi di accoppiamento rimangono morfologicamente identici ma presentano differenze fisiologiche. In P. aurelia, ci sono 14 singeni e 28 tipi di accoppiamento, mentre in P. caudatum, ci sono 16 singeni e 32 tipi di accoppiamento. Si è osservato che di solito i paramecia non si coniugano né con i membri del proprio tipo di accoppiamento né con le altre varietà, ma solo con il secondo tipo di accoppiamento della propria varietà.

Fattori che inducono la coniugazione:
I fattori che inducono la coniugazione variano da specie a specie, ma alcuni di essi sono riportati di seguito:
1. La coniugazione avviene di solito in condizioni di vita sfavorevoli; si dice che la fame o la carenza di cibo e una particolare dieta batterica o alcune sostanze chimiche inducano il processo di coniugazione in alcune specie di Paramecium.
2. La coniugazione avviene dopo circa 300 generazioni asessuali di fissione binaria, o si alterna con la fissione binaria a lunghi intervalli per ringiovanire il clone morente, cioè, si verifica negli individui che devono essere passati attraverso un numero desiderabile di generazioni asessuali, detto periodo di immaturità, e poi diventano sessualmente maturi per coniugarsi.
3. La coniugazione si verifica quando c’è un cambiamento nella condizione fisiologica dei parameci, quindi si verifica tra tali individui che sono un po’ più piccoli in termini di dimensioni (210 micron di lunghezza) e sono in uno stadio che può essere considerato come un periodo di vecchiaia malsana; i parameci di questa condizione moriranno se non si permette loro di coniugare.
4. Si dice che il buio improvviso in condizioni di luce e le basse temperature inducano il processo di coniugazione in alcune specie.
5. La coniugazione non avviene durante la notte o l’oscurità; inizia al mattino presto e continua fino al pomeriggio.
6. Si dice che una sostanza proteica nelle ciglia degli individui di tipo accoppiamento induca la coniugazione.
Processo di coniugazione:
Il processo di coniugazione differisce nelle diverse specie di Paramecium, ma il racconto qui sotto riportato si riferisce al processo di coniugazione di P. caudatum (Fig. 20.21).
Nella coniugazione due Paramecium caudatum (detti preconiuganti) dei tipi di accoppiamento opposti della stessa varietà si avvicinano con le loro superfici ventrali e si uniscono attraverso i loro solchi orali; le loro ciglia producono una sostanza sulla superficie del corpo che causa l’adesione dei due parameci coniuganti.
Cessano di nutrirsi e il loro apparato dei solchi orali scompare. La pellicola e l’ectoplasma, nel punto di contatto, di entrambi si rompono, e un ponte protoplasmatico si forma tra i due animali. Ora, questi individui sono chiamati coniuganti.
In questa condizione, la coppia coniugante nuota attivamente e contemporaneamente una serie di cambiamenti nucleari hanno luogo in ogni coniugante come descritto di seguito:
Il macronucleo comincia a disintegrarsi, diventa di consistenza molle e forma una complessa matassa attorcigliata, durante la seconda metà del periodo di coniugazione infine scomparirà venendo assorbito nel citoplasma. Il micronucleo di ogni coniugato si divide due volte, una delle quali è una divisione di riduzione.
Così, quattro micronuclei figlia aploide sono prodotti in ogni coniugante. Tre di questi quattro micronuclei degenerano in ciascuno, così che ne rimane solo uno.
Il micronucleo rimanente di ogni coniugante si divide mitoticamente in due pronuclei o nuclei gametici disuguali che formano un pro-nucleo femminile stazionario più grande e un pro-nucleo maschile migratorio più piccolo e attivo.
Il pro-nucleo migratorio di un coniugato attraversa il ponte protoplasmatico e si fonde con il pro-nucleo stazionario dell’altro coniugato per formare un synkaryon o nucleo di coniugazione in cui il numero diploide di cromosomi è ripristinato e c’è stato uno scambio di materiale ereditario.
Il processo è stato paragonato alla fecondazione negli animali superiori, ma questa non è una fecondazione perché non sono coinvolti gameti. I coniuganti ora (dopo circa 12-48 ore) si separano e sono chiamati ex coniuganti. Il sincario di ogni ex coniugato si divide tre volte per formare otto micronuclei in ogni ex coniugato.
Quattro degli otto micronuclei si ingrandiscono e diventano macronuclei, e tre degli altri quattro micronuclei scompaiono.
Il micronucleo rimanente si divide e allo stesso tempo l’ex coniugato si divide per fissione binaria in due cellule, ciascuna con due macronuclei e un micronucleo. Le cellule e i loro micronuclei si dividono una seconda volta per formare quattro paramecia da ogni ex coniugato, in modo che ognuno abbia un macronucleo e un micronucleo.
Il nuovo macronucleo, come anche il micronucleo, sono stati fatti di materiale nuovo. Questi nuovi nuclei contengono probabilmente un nuovo e diverso potenziale che si riflette negli individui sani.
Significato della coniugazione:
Un clone si estingue se non si verifica la riorganizzazione nucleare, ma il clone può essere ringiovanito per riacquistare il suo antico vigore attraverso il riarrangiamento nucleare, questa riorganizzazione nucleare è portata dalla coniugazione, quindi, la coniugazione è essenziale per continuare la fissione binaria.
Il significato della coniugazione è stato riassunto qui sotto:
1. La coniugazione serve come un processo di ringiovanimento e riorganizzazione attraverso il quale viene ripristinata la vitalità della razza. Se la coniugazione non avviene per lunghi periodi, i parameci si indeboliscono e muoiono. (La pretesa di Woodruffs di mantenere i parameci sani per 22.000 generazioni senza coniugazione è smentita da Sonneborn, perché ha dimostrato che tutti i parameci di Woodruffs appartenevano allo stesso tipo di accoppiamento).
2. Non c’è distinzione di sesso nei coniugi, anche se solo i parameci di due diversi tipi di accoppiamento della stessa varietà si coniugano.
3. Non c’è distinzione di sesso, ma il pro-nucleo migratorio attivo è considerato maschio e il pro-nucleo stazionario è considerato femmina.
4. La coniugazione è solo un’unione temporanea, non c’è fusione del citoplasma e non viene prodotto uno zigote, ma il nucleo di ogni ex coniugato contiene materiale ereditario di due individui coniugati.
5. La coniugazione porta alla sostituzione del macronucleo con materiale proveniente dal sincario, questo è un evento di fondamentale importanza. Nella fissione binaria i cromosomi del macronucleo erano distribuiti a caso alle cellule figlie, la continua fissione binaria aveva reso il clone debole con alcune anomalie strutturali.
La coniugazione porta alla formazione del corretto numero di cromosomi nel macronucleo, in modo che la razza si rinnovi nel vigore. Il ruolo del micronucleo è quello di ripristinare un complesso cromosomico e genico equilibrato.
Comportamento aberrante nella riproduzione nel Paramecium Caudatum:
Paramecium Caudatum mostra alcune variazioni nel suo comportamento nucleare durante la fissione e la coniugazione, queste deviazioni sono endomissia, autogamia, citogamia ed emissia. Nei primi tre processi avviene la ricombinazione genetica e dal micronucleo si forma un nuovo macronucleo.
(i) Endomissia:
Woodruff e Erdmann (1914) riportarono per primi un nuovo processo di riorganizzazione nucleare, l’endomissia (Gr., endon = dentro; mixis = mescolanza) in Paramecium aurelia, una specie bimicronucleata (Fig. 20.22). Questo processo è stato descritto come un processo che si verifica periodicamente in cui un nuovo apparato macronucleare è prodotto senza formazione di sincario. L’endomissia si verifica all’interno di un singolo individuo.
Secondo Woodruff e Erdmann, il macronucleo degenera e i micronuclei si dividono due volte per formare otto micronuclei. Sei micronuclei degenerano e ne rimangono due. Con solo due micronuclei rimasti, l’animale si divide per fissione in due cellule, ciascuna con un micronucleo.
Il micronucleo di ogni cellula figlia si divide due volte per formare quattro micronuclei. Due micronuclei si ingrandiscono per formare macronuclei. L’animale e i suoi micronuclei si dividono in modo da formare due individui figli, ciascuno con un macronucleo e due micronuclei.
L’endomissia si verifica in quella varietà di P. aurelia che non si coniuga, quindi, l’effetto dell’endomissia può essere lo stesso di quello della coniugazione poiché entrambi i processi portano alla sostituzione del macronucleo con materiale del micronucleo, ed entrambi i processi ringiovaniscono la vitalità della razza.
Ma i due processi differiscono perché non c’è fusione di pronuclei nell’endomissia; l’endomissia può essere paragonata alla partenogenesi.
Tuttavia, alcuni operatori hanno sostenuto con buone ragioni che l’endomissia non è un processo valido ed è stato descritto a causa di un’osservazione errata. Con ogni probabilità l’endomissia non ha luogo e può essere solo un caso specializzato di autogamia.

In seguito, Erdmamm e Woodruff (1916) riportarono l’endomissia in Paramecium caudatum. Diller, tuttavia, non crede nella validità di questo processo e ritiene che Erdmann e Woodruff abbiano semplicemente combinato le fasi di emixia e autogamia in un unico schema, l’endomissia.
(ii) Autogamia:
Diller (1934, 1936) e Sonneborn (1950) hanno descritto un processo di autofecondazione o autogamia che si verifica in un singolo individuo in Paramecium aurelia (Fig. 20.23). Egli ha riferito che nell’autogamia tre divisioni micro-nucleari (pregamiche), con maturazione, producono i nuclei gametici (pronuclei).
Durante l’autogamia in P. aurelia, i due micronuclei si dividono due volte (una volta meioticamente) per formare otto micronuclei, sei dei quali degenerano. Nel frattempo il macronucleo cresce in una massa simile a una matassa che si rompe in pezzi più tardi per essere assorbita nel citoplasma. Due degli otto micronuclei, come pronuclei, entrano in un cono protoplasmatico rigonfio vicino alla bocca della cellula.
I due pronuclei si fondono per formare il sincario. Il sincario si divide due volte per formare quattro micronuclei. Due micronuclei diventano macronuclei. Il Paramecium e i suoi micronuclei si dividono per formare due individui figli, ciascuno con un macronucleo e due micronuclei. Questo processo si completa in circa due giorni.
L’autogamia porta al ringiovanimento della razza.
Somiglia alla coniugazione in quanto il nuovo macronucleo è formato da materiale proveniente dal micronucleo, nel nuovo macronucleo viene ripristinato il numero corretto di cromosomi; e anche nel fatto che avviene la fusione di due pronuclei. Ma l’autogamia differisce dalla coniugazione perché solo un individuo prende parte all’autogamia e fornisce entrambi i pronuclei, è una sorta di autofecondazione.
(iii) Emixis:
Diller (1936) ha riportato l’emixis in Paramecium aurelia (Fig. 20.24). L’emixis è principalmente un processo di frammentazione e divisione macro-nucleare senza alcuna attività micro-nucleare insolita. Diller ha classificato l’emissi in quattro tipi, cioè A, B, C, e D come mostrato nella Fig. 20.24 in P. aurelia, ma ha anche incontrato tutti i tipi in colture di massa di P. caudatum e P. multimicronucleatum.
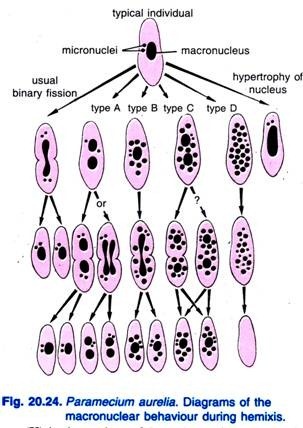
Il tipo A è la forma più semplice di emixis caratterizzata da una divisione del macronucleo in due o più parti. Questa divisione non è sincronizzata con la divisione micro-nucleare.
Il tipo B è caratterizzato dall’estrusione di una a 20 o più sfere di cromatina dal macronucleo nel citoplasma.
Il tipo C è caratterizzato dalla scissione simultanea del macronucleo in due o più porzioni principali e dall’estrusione di sfere macronucleari nel citoplasma.
Il tipo D è considerato rappresentare condizioni patologiche in cui il macronucleo subisce una frammentazione completa in sfere di cromatina che alla fine scompaiono dalla cellula. I micronuclei generalmente scompaiono prima della dissoluzione del macronucleo.
(iv) Citogamia:
Wichterman (1939) ha riportato un altro processo sessuale nel Paramecium caudatum, che ha chiamato citogamia. Nella citogamia, non c’è scambio nucleare. In questo processo, due individui si uniscono per le loro superfici ventrali, ma la pellicola dei due individui non si rompe.
Il micronucleo di ogni individuo si divide tre volte per formare otto micronuclei, sei dei quali si disintegrano in ogni individuo. I due micronuclei rimanenti si fondono per formare un sincario in ogni cellula. Gli animali ora si separano.
La citogamia differisce dall’autogamia in quanto ci sono due animali in contatto tra loro, ma assomiglia all’autogamia e alla coniugazione nella fusione di due pronuclei. La citogamia differisce dalla coniugazione in quanto non c’è scambio nucleare tra i due animali che si uniscono.
Alcune particelle citoplasmatiche riportate nel Paramecium Caudatum:
(i) Particelle Kappa:
Nel 1938, T.M. Sonneborn riferì che alcune razze (note come killer o ceppo killer) di Paramecium producono una sostanza velenosa, chiamata paramecina che è letale per altri individui chiamati sensibili. La paramecina è solubile in acqua, diffusibile e dipende per la sua produzione da alcune particelle situate nel citoplasma del Paramecium (ceppo killer).
Queste particelle sono chiamate particelle kappa. Le particelle kappa hanno DNA e RNA. Un Paramecium killer può contenere centinaia di particelle kappa. Lo studio dettagliato di queste particelle ha rivelato che un gene dominante (K) nel nucleo del Paramecium è necessario per le particelle kappa per esistere, moltiplicarsi e produrre paramecina.
(ii) mµ, particelle:
R.W. Siegel (1952) ha riportato un altro tipo di particelle killer nel citoplasma di alcuni Paramecium. Un Paramecio con particelle mp è chiamato mate killer perché quando si coniuga con un Paramecio senza particelle mµ chiamato mate sensitive, allora uccide quest’ultimo. Le particelle mp sono anche composte da DNA, RNA, ecc.
Queste particelle esistono solo in quei parameci il cui micronucleo contiene almeno un gene dominante di una delle due coppie di geni cromosomici non collegati (M1 e M2).
(iii) Particelle Pi:
Si suppone che queste particelle siano la forma mutante delle particelle kappa ma non producono alcun tipo di sostanza velenosa.
(iv) Particelle Lambda:
Queste particelle sono segnalate nei paramecia killer e si dice che producano qualche sostanza responsabile di causare la lisi o la disintegrazione dei paramecia sensibili, cioè che non la possiedono.