Gametes och befruktning översikt
Könslig reproduktion avser de sexuella könscellerna (gameterna) som produceras av föräldrarna, genom kombinationen av bisexuella könsceller (t.ex. spermie- och äggceller), som blir till det befruktade ägget, och sedan utvecklas det befruktade ägget till en ny individ, så kallad sexuell reproduktion. Omfattande variation i genkombinationer vid sexuell reproduktion kan öka avkommans förmåga att anpassa sig till det naturliga urvalet. Slumpmässigt kombinerade gener i avkommor från sexuell reproduktion kan vara till nytta för arten eller inte, men ökar åtminstone chansen att några få individer överlever i en oförutsägbar och ständigt föränderlig miljö, vilket gynnar arten. Sexuell reproduktion kan också främja spridningen av gynnsamma mutationer i populationen. Om två individer i en art har gynnsamma mutationer på olika platser, i den asexuella reproduktionspopulationen, kommer de två mutanterna att konkurrera tills en eliminering, det är omöjligt att behålla båda gynnsamma mutationer samtidigt. I sexuellt reproducerande populationer kan dock dessa två gynnsamma mutationer genom parning och rekombination samtidigt komma in i arvsmassan hos samma individ och samtidigt spridas i populationen. Av ovanstående skäl påskyndar den sexuella reproduktionen evolutionsprocessen. Under de mer än 3 miljarder år som den biologiska utvecklingen på jorden har pågått, har livets första två miljarder år förblivit i det asexuella reproduktionsstadiet, och utvecklingen har varit långsam, och utvecklingstakten har accelererat betydligt sedan den senaste 1 miljard åren. Förutom förändringar i jordens miljö (t.ex. uppkomsten av den syrehaltiga atmosfären etc.) är förekomsten och utvecklingen av sexuell reproduktion också en viktig orsak.
Gametes
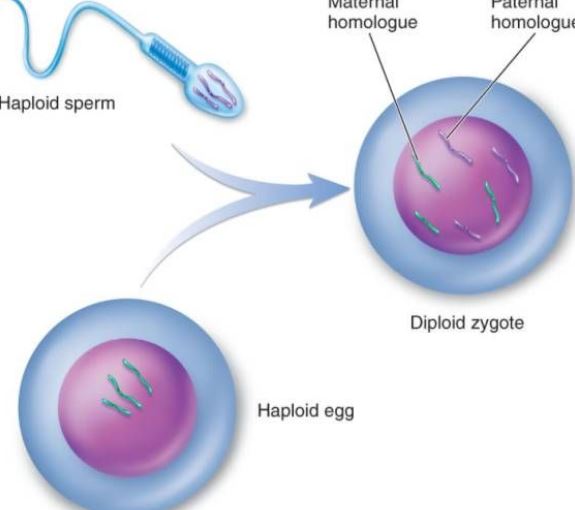
Gametes avser mogna celler som produceras av fortplantningssystemet när organismen reproducerar sig sexuellt, så kallade könsceller. Gameterna delas in i hanliga könsceller och kvinnliga könsceller. De kvinnliga könscellerna hos djur och växter brukar kallas äggceller och de manliga könscellerna kallas spermier. Spermierna är ganska små, men de kan röra sig och gå in i äggcellen i en skorpionform. Äggcellen är ganska stor och simmar inte. Till exempel är äggcellens volym hos sjöborren 10 000 gånger större än spermiecellernas. Även om de manliga och kvinnliga könscellerna skiljer sig åt i volym är det kärn-DNA som de förser avkomman med lika mycket, det vill säga var och en förser den med en uppsättning genomer. På grund av äggcellernas stora storlek tillhandahålls dock i princip dottercellernas cytoplasmatiska struktur och cytoplasmatiska DNA av äggcellerna. Gameterna spelar en viktig roll i biologiska beräkningar. Genom genetiska kartor kan genflödet och mönstret för avkommans genotyper tydligt observeras. Antalet könsceller i den första generationen hybrider och de relativa egenskaperna hos hybriderna är 2nth power (n är logaritmen av de relativa egenskaperna). När en levande kropp bildar en gamet separeras de parade genetiska faktorerna från varandra och går in i olika gameter. Gameterna innehåller endast en av varje par genetiska faktorer. Vid befruktning är kombinationen av manliga och kvinnliga gameter slumpmässig. På grund av det släkte av könsceller som bildas genom meiosen, mångfalden i kromosomsammansättningen, skillnaden i genetiskt material hos olika könsceller och slumpmässigheten i kombinationen av äggceller och spermier i befruktningsprocessen måste avkomman av samma förälder vara varierande.
Fertilisering
Fertilisering är den process genom vilken ägg och spermier smälter samman till en zygot. Den är ett grundläggande inslag i den sexuella reproduktionen och är allestädes närvarande i djur- och växtvärlden, men den är ofta den mest omnämnda. Djurens befruktning på cellnivå omfattar befruktningsprocessen tre huvudfaser: aktivering av ägget, reglering och amfoterisk pronukleär fusion. Aktiveringen kan betraktas som startpunkten för ontogenin, som huvudsakligen manifesteras av förändringar i oozingmembranets genomsläpplighet, ecclesia av kortikala granuler, bildandet av det befruktade membranet osv. Justering sker efter aktivering, vilket är det första steget för att säkerställa normal delning av befruktade ägg. Förändringen av den prokaryotiska kärnan säkerställer arvet från föräldrarna och återställer det diploida. Befruktningen initierar inte bara replikationen av DNA utan aktiverar också den genetiska informationen såsom mRNA och rRNA i ägget för att syntetisera det protein som krävs för embryoutvecklingen. Efter 6 till 7 dagar efter befruktningen försvinner blastocysten zona pellucida och begravs gradvis och täcks av livmoderslemhinnan, vilket kallas implantation av det befruktade ägget. Spermierna hos ett djur har inte någon tydlig kemotaxis som spermier hos lägre växter, t.ex. bryofyter, utan når snarare äggets närhet genom aktiv rörelse eller genom de genitala epitelcellernas ciliära rörelser. Spermakapacitering: Det är känt att när många spermier från däggdjur passerar genom de kvinnliga könsorganen eller genom cumulus, avlägsnas det främmande protein som kapslar in spermierna, och de fysiska och biologiska egenskaperna hos spermiernas plasmamembran förändras, vilket gör det möjligt för spermierna att få energi och delta i befruktningsprocessen. När den kapabiliserade spermien från däggdjur kommer i kontakt med äggets membran eller zona pellucida binder den specifikt till ett glykoprotein på äggmembranet, vilket stimulerar spermien att producera energi. Akrosomreaktionen hjälper spermierna att korsa äggmembranet ytterligare. På sjöborreägget är spermiernas akrosomreaktion en viss polysackaridsubstans i perimembranfilmen. De flesta ägg har äggmembran i periferin, och tjockleken på de olika äggmembranen varierar. Huvudkomponenten är mucin eller mukopolysackarid. Endast ett fåtal är nakna ägg, t.ex. ägg från en coelenterat. Acrosomreaktion: När spermierna passerar genom äggmembranet sker en bindningsprocess. Den förstnämnda är löst fäst, fri från yttre temperaturstörningar och har ingen specificitet. Under vidhäftningsperioden omvandlas protoplastproteinet på det akrosomala membranet till akrosomprotein, och akrosomproteinet påskyndar spermiernas passage genom äggmembranet; Det är en stark kombination, kan störas av låg temperatur och har en specificitet. Ett protein som binder specifikt till äggglykoproteinet har isolerats på plasmamembranet hos spermier från sjöborrar, ett s.k. bindningsprotein, med en molekylvikt på ca 30 000. De sekundära äggcellerna i mitten av den andra mognadsdivisionen under ägglossningen, tillsammans med den omgivande zona pellucida och strålningskronan, går snabbt in i äggledarens ampulla genom bukhålan på grund av svängningen av cilierna i äggledarens epitelceller och sammandragningen av muskelskiktet. Om äggcellen misslyckas med att möta spermier börjar den vanligtvis degenerera inom 12-24 timmar. När spermierna väl är i kontakt med ägget genomgår själva ägget en rad aktiveringsförändringar. På däggdjursägg kännetecknas den av den kortikala reaktionen, äggledarmembranreaktionen och zona pellucida-reaktionen, som kan blockera flerbefruktningen och stimulera den fortsatta utvecklingen av ägg. Den kortikala reaktionen inträffar vid tidpunkten för spermiecellernas sammansmältning. Från fusionspunkten spricker kortikalpartiklarna och innehållet utlöses, vilket sprider kortikalen i hela ägget. Eimembranreaktionen är en rekombinationsprocess av ägget och det kortikala granulathöljet. Zona pellucida-reaktionen är en process där det kortikala utflödet och zona pellucida från ett befruktat membran, äggmembranet separeras från plasmamembranet, spermiereceptorn i zona pellucida försvinner och zona pellucida hårdnar. Endast spermier som har en akrosomreaktion kan smälta samman med ägget. Under inverkan av akrosin passerar spermierna genom den radiella kronan och interagerar med spermiereceptorns glykoproteinmolekyl ZP3 på zona pellucida, vilket gör det möjligt för spermierna att frigöra det akrosomala enzymet och gå in i perivitellin genom zona pellucida. I början av befruktningen är höljet av den ekvatoriella delen av det mänskliga spermiehuvudet i kontakt med äggcellsmembranet, och därefter kommer spermiernas kärna och cytoplasma in i ägget. Efter att spermien trängt in i ägget släpper de kortikala partiklarna i äggets grunda cytoplasma omedelbart sitt innehåll till membranets periferi. Samtidigt, när de kortikala granulernas membran smälter samman med äggcellerna, ökar den negativa ytladdningen på cellytan, vilket undertrycker fusionen av spermiernas plasmamembran med äggmembranet, vilket kallas för en kortikal reaktion. Zona pellucidas strukturförändringar kallas zona pellucida-reaktionen. Vid denna tidpunkt minskar zona pellucidas förmåga att binda spermier, vilket förhindrar förekomsten av polyspermier och säkerställer de biologiska egenskaperna hos mänsklig monospermi. Förutom att förstöra eller inaktivera den receptor som är bunden till spermierna i zona pellucida kan innehållet i de kortikala granulerna också öka tvärbindningen mellan peptidkedjorna i zona pellucida, försvaga känsligheten för akrosomproteaset och förhindra spermiernas penetration. Även om flera spermier passerar genom zona pellucida är det bara en spermie som kommer in i äggcellen för att befrukta den. I en onormal situation är det två spermier som deltar i befruktningen, nämligen dubbel spermiebefruktning. Två spermier går samtidigt in i ägget och bildar embryon av triploida celler, som alla aborteras eller dör strax efter födseln. Efter att spermierna trängt in i ägget genomför ägget snabbt den andra mogna delningen. Vid denna tidpunkt kallas spermiernas och äggets kärnor för manlig pronukleus respektive kvinnlig pronukleus. De två pronukleerna närmar sig gradvis varandra, kärnmembranet försvinner och kromosomerna smälter samman för att bilda ett diploid befruktat ägg. När spermierna smälter samman kan man se att mikrovilli på äggets yta omger spermierna, som kan vara orienterade; sedan smälter äggledarmembranet samman med plasmamembranet i spermiernas bakre del av akrosomet.
Referens
- Yanagimachi R. Germ cells and fertilization: why I studied these topics and what I learned along the path of my study. Andrologi. 2014, 2(6):787-93.
- Henshaw J M, Marshall D J, Jennions M D, et al. Local gamete competition explains sex allocation and fertilization strategies in the sea. American Naturalist. 2014, 184(2):32.
- Mori T, Igawa T. Gamete attachment process revealed in blooming plant fertilization. Plant Signaling & Behavior. 2014, 9(12):977715.
- Hédouin L, Pilon R, Puisay A. Hyposalinitetsstress äventyrar befruktningen av gameter mer än överlevnaden hos koralllarver. Marin miljöforskning. 2015, 104:1-9.
Luo J, Mcginnis L K, Carlton C, et al. PTK2b-funktion under befruktning av musens oocyt. Biochem Biophys Res Commun. 2014, 450(3):1212-1217.