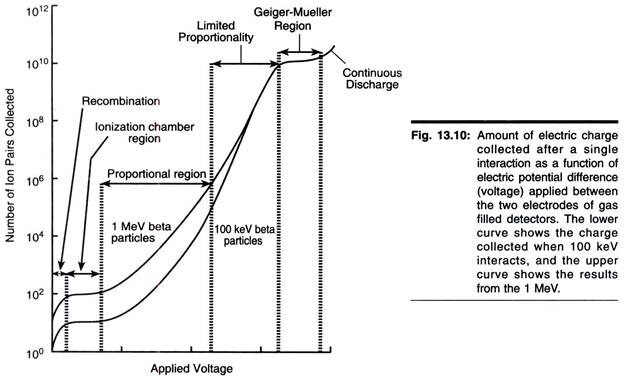
ADVERTISEMENTS:
I den här artikeln kommer vi att diskutera Paramecium caudatum:- 1. Habitat, livsmiljö och kultur av Paramecium caudatum 2. Struktur av Paramecium Caudatum 3. Rörelse 4. Näring 5. Respiration och utsöndring 6. Osmoreglering 7. Beteende 8. Reproduktion 9. Avvikande beteende vid reproduktion 10. Några cytoplasmatiska partiklar.
Innehåll:
- Habitus, Habitat och odling av Paramecium caudatum
- Struktur hos Paramecium caudatum
- Lokomotion hos Paramecium caudatum
- Näring hos Paramecium caudatum
- Respiration och utsöndring hos Paramecium caudatum
- Osmoregulering hos Paramecium Caudatum
- Beteende hos Paramecium Caudatum
- Förökning hos Paramecium Caudatum
- Avvikande beteende vid reproduktion hos Paramecium Caudatum
- En del cytoplasmatiska partiklar rapporteras hos Paramecium Caudatum
- Habitat, Habitat och odling av Paramecium caudatum:
- Kultur av Paramecium:
- Struktur hos Paramecium caudatum:
- (i) Storlek och form hos Paramecium caudatum:
- (ii) Pellicle:
- (iii) Cili:
- (iv) Infra-ciliärt system:
- (v) Munspår och cytopyge:
- (vi) Cytoplasma:
- (vii) Ektoplasma:
- (viii) Trikocystor:
- (ix) Neuromotorium och tillhörande fibriller:
- (x) Endoplasma:
- (xi) Kontraktila vakuoler:
- (xii) Livsmedelsvakuoler:
- Förflyttning hos Paramecium Caudatum:
- (i) Metabolism eller kroppsrörelser:
- (ii) Ciliär förflyttning:
- Näring hos Paramecium Caudatum:
- Föda mekanism:
- Förtäring och uppslutning hos Paramecium Caudatum:
- Respiration och exkretion hos Paramecium Caudatum:
- Paramecium caudatums beteende:
- (i) Reaktioner på kontakt (Thigmotaxis):
- (ii) Reaktioner på kemikalier (Chemotaxis):
- (iii) Reaktioner på temperatur (Thermotaxis):
- (iv) Reaktioner på ljus (Phototaxis):
- (v) Reaktioner på elektrisk ström (Galvanotaxis):
- (vi) Reaktioner på vattenström (Rheotaxis):
- (vii) Reaktioner på gravitation (Geotaxis):
- Reproduktion hos Paramecium Caudatam:
- (i) Tvärgående binär klyvning:
- (ii) Konjugering:
- Aberrant beteende vid reproduktion hos Paramecium Caudatum:
- (i) Endomixis:
- (ii) Autogami:
- (iii) Hemixis:
- (iv) Cytogami:
- Vissa cytoplasmatiska partiklar som rapporterats i Paramecium caudatum:
- (i) Kappapartiklar:
- (ii) mµ, partiklar:
- (iii) Pi-partiklar:
- (iv) Lambdapartiklar:
Habitat, Habitat och odling av Paramecium caudatum:
Paramecium caudatum (Gr., paramekes = avlång; L., caudata = svans) är vanligt förekommande i sötvattendammar, bassänger, diken, bäckar, sjöar, reservoarer och floder. Den förekommer särskilt rikligt i stillastående dammar som är rika på rötande material, i organiska uppblandningar och i avloppsvatten. Paramecium caudatum är en frilevande organism och arten är spridd över hela världen.
Kultur av Paramecium:
RÅDGÅNGAR:
Tag undervattens ogräs från en damm och lägg det i en burk med destillerat vatten, täck burken och låt den ruttna; svärmar av Paramecia kommer att dyka upp inom några dagar. Koka nu hö i vatten, dekantera infusionen och tillsätt några vetekorn och låt den stå tills den är grumlig av bakterier.
Överför Paramecia från den första burken till denna vätska där de snabbt kommer att föröka sig. Enbart höinfusioner ger Paramecia som visar närvaro av cystor, och cystor som liknar sandkorn har rapporterats, men det finns inga bevis för att Paramecia bildar cystor, eftersom de aldrig har bekräftats.
Struktur hos Paramecium caudatum:
(i) Storlek och form hos Paramecium caudatum:
Paramecium caudatum (Fig. 20.1) är en mikroskopisk organism och är synlig för blotta ögat som en mycket liten långsträckt kropp.
Åtgärdsförslag:
Den framstår som ljusgrå eller vit och mäter vanligen mellan 170 och 290 mikrometer i längd och kan nå en längd på upp till 300-350 mikrometer. P. caudatum ser ut som sulan på en toffel eller sko, därav är djuret allmänt känt som toffeldjur. Den är fyra gånger så lång som bred och något cylindrisk med tydligt olika ändar.
Den framåtriktade främre delen är slank med en trubbig eller rundad ände, medan den bakre änden är något spetsig eller kägelformad. Organismens bredaste del ligger strax under mitten. Djurets kropp är asymmetrisk till formen och visar en väldefinierad oral eller ventral yta och en aboral eller dorsal yta.
(ii) Pellicle:
Kroppen är täckt av en tunn, dubbelskiktad, elastisk och fast pellicle av gelatin. Pelliceln håller djurets form men är tillräckligt elastisk för att tillåta sammandragningar. Pelliceln har dubbla membran, det yttre membranet är kontinuerligt med cilierna och det inre membranet med ektoplasman. I mikroskopets högre förstoring visar pelliceln rektangulära eller hexagonala fördjupningar på sin yta.

Detta arrangemang finns bevarat på Parameciums dorsala yta, men på den ventrala ytan konvergerar åsarna fram och bakåt mot en preoral och postoral öppning. Varje sexkantig fördjupning är perforerad av en central öppning genom vilken ett enda cilium kommer ut. De sexkantiga fördjupningarnas främre och bakre marginaler är försedda med öppningar för trikocystor.
Ehret och Powers (1957) elektronmikroskopiska undersökning av pellicle (fig. 20.2) har visat att de hexagonala fördjupningarna motsvarar regelbundna serier av håligheter, alveolerna. Alla alveoler bildar tillsammans ett kontinuerligt alveolärt skikt, som avgränsas av ett yttre alveolärt membran och ett inre alveolärt membran.
Det yttre lagret ligger i nära kontakt under det yttre cellmembranet. Därför omfattar pellicle det yttre cellmembranet, det yttre alveolarmembranet och det inre alveolarmembranet.
(iii) Cili:
Hela kroppen är täckt av många, små, hårliknande utsprång som kallas cilier. Ciliorna förekommer i längsgående rader över hela kroppen, detta tillstånd är känt som holotrikt, där kroppens cilior är lika stora. Cilior har samma struktur som flageller, de har ett yttre protoplasmatiskt hölje eller plasmamembran med nio dubbla längsgående fibriller i en perifer ring. I vissa cilier är de nio yttre fibrillerna inte parade.
ÅTERKÄNNANDE:
Det finns två centrala longitudinella fibriller som är tunnare än de yttre fibrillerna. Varje cilium utgår från en basal granula eller kinetosom. De nio paren av perifera fibriller smälter samman och bildar kinetosomens vägg, kinetosomen är alltså ett rör som antingen är öppet eller stängt i sin nedre ände, de två centrala fibrillerna slutar på pellicelnivå hos de flesta ciliater.
Utgående från kinetosomen är en tunn rhizoplast som inte ansluter till kärnan. Många Metazoa har också cilier, deras struktur är densamma, förutom att den basala granulen är annorlunda och den har fina filament eller rotfibrer som sträcker sig ner i cytoplasman. Men cilierna skiljer sig från flagellerna genom att de i allmänhet är talrikare och kortare i storlek.
Ciliaturen kan lämpligen delas in i kropps- eller somatiska cilier som finns på kroppsytan, och i orala cilier som är förknippade med munregionen. Kroppsciliorna är lika stora men de är längre i den bakre änden, därav namnet caudatum. Ciliorna är organeller som används för att förflytta sig och samla in föda. De fungerar också som sensoriska receptorer och registrerar stimuli från den yttre miljön.

Ultrastruktur av cilier:
Cilierna och flagellerna har en fibrillär sammansättning. Vid basen har ciliumet en diameter på cirka 0,2 mikrometer eller 2 000 A0 som kan vara upp till 10 mikrometer över cellytan. Cilierna avgränsas av ett enhetsmembran med en tjocklek på 90 A0 som liknar och förblir kontinuerligt med plasmamembranet. Det avgränsade utrymmet för cilierna innehåller en vattnig substans som kallas matris.
I matrisen förblir inbäddade elva längsgående fibriller eller mikrotubuli. Av de elva fibrillerna är två placerade i centrum, medan de övriga nio fibrillerna förblir arrangerade perifert runt de centrala fibrillerna. Var och en av de nio yttre fibrillerna har en diameter på 360 A0 och består av två subfibriller med en diameter på 180-250 A0.
Dessa subfibriller betecknas som subfibrill A och subfibrill B.
ADVERTISEMENTS:
Subfibrillen A är något större än subfibrillen B. Subfibrillen A ger ut två tjocka utsprång eller armar från sin ena sida. Armarna från subfibrill A i alla de yttre fibrillerna förblir riktade i riktning medurs. Dessutom ligger subfibrill A närmare ciliumets centrum än subfibrill B. Båda subfibrillerna har en gemensam vägg med en tjocklek på 50A0.
De två centrala fibrillerna har inte parvisa subfibriller som de perifera nio fibrillerna utan var och en innehåller endast en enda tubuli. Varje central fibrill har en diameter på cirka 250A0 och består av en 60A0 tjock vägg.
Båda de centrala fibrillerna förblir åtskilda av ett utrymme på 350A0 och förblir omslutna av ett gemensamt hölje. Gibbnos (1967) har rapporterat att de centrala fibrillernas hölje ger ut nio radiellt orienterade länkar eller ekrar till varje underfibrill A.

Den högupplösta elektronmikroskopin har avslöjat att var och en av de perifera och centrala fibrillerna hos cilierna och flagellerna består av tio till tolv filament med en tjocklek på 40A0. Varje filament är pärlformigt. Varje pärla förblir arrangerad i nät av 40 x 50A0 i planet av tubulära väggar. Dessa pärlor anses vara den grundläggande underenheten i tubulstrukturen.
(iv) Infra-ciliärt system:
Infraciliärsystemet är beläget strax under de pellikulära alveolerna. Det består av kinetosomen eller basalkroppen och kinetodesma. Ciliorna uppstår från kinetosomerna och från varje kinetosom uppstår en känslig cytoplasmatisk fibrill som kallas kinetodesma, (fig. 20.2).
Under pelliceln ligger något till höger, men förenat med alla kinetosomer i en längsgående rad, ett längsgående knippe av flera kinetodesmata, ett kinetodesmata från varje kinetosom sträcker sig ett stycke framåt i sitt eget knippe av kinetodesmata.
En longitudinell rad av kinetosomer med sina kinetodesmata bildar en longitudinell enhet som kallas kinety. Alla kineties eller kinetia utgör det infraciliära systemet hos en ciliat. Kinetierna ligger i cortexen under pelliculan, deras antal är ganska konstant för varje ciliat.
Infraciliärsystemet kontrollerar och samordnar ciliernas rörelser, och det åstadkommer bildandet av organeller vid celldelning, t.ex. bildar vissa kinetia munnen. Vid binär klyvning hos ciliater skärs kinetierna tvärs över i två, var och en går till en dottercell, detta kallas perikinetal klyvning.
(v) Munspår och cytopyge:
På den ventrolaterala sidan finns en stor snedställd, grund fördjupning som kallas munspår eller peristom och som ger djuret ett asymmetriskt utseende. Den löper snett bakåt från ena sidan (vanligtvis från vänster till höger men i vissa fall från höger till vänster) och slutar en bit bakom mellankroppen. Munspåret mynnar ut i en kort konisk trattformad fördjupning som kallas vestibulum.
ADVERTIVERINGAR:
Vestibulum leder direkt in i den fasta, ovalformade öppning som kallas cytostome (mun). Från cytostomen sträcker sig det breda cytofarynx direkt mot kroppens mitt. Cytopharynx svänger sedan skarpt mot baksidan för att bli den smala, avsmalnande matstrupen (oesophagus).
Sålunda är matstrupen ungefär parallell med kroppsyta hos Paramecium utom vid dess bakre ände. Här vänder sig matstrupen återigen mot djurets centrum för att leda in i den bildande matvakuolen.
Cytopygen (även kallad cellanus eller analfläck eller cytoprokt) ligger på kroppens ventrala yta nästan vertikalt bakom cytostomen eller munnen. Osmälta matpartiklar elimineras genom cytopygen. Cytopharynx ciliation är mycket komplicerad.
Gelei (1934) rapporterade förekomsten av fyra rader och Lund (1941) observerade minst fyra rader. En struktur som kallas penniculus finns på cytofarynxens vänstra vägg och spiralformas i ungefär 90 grader så att dess bakre ände befinner sig på den orala (ventrala) ytan av matstrupen.
Enligt Lund består penniculus av åtta rader av cilier arrangerade i två tätt placerade block av varje. Ett liknande band som består av fyra rader långa cilier som är mindre kompakta än i penniculus kallas quadrulus. Det går i en spiral längs buccalhålans dorsala vägg och slutar nära penniculus.
Penniculus och quadrulus har felaktigt kallats för ett böljande membran av vissa arbetare. Quadrulus och penniculus kontrollerar passagen av föda. Man vet inte hur cilierna fungerar, troligen drar deras fibriller ihop sig på ett rytmiskt sätt som orsakar böjning. Gelei (1925) påpekade att penniculus funktion är att tvinga in födoämnen i kroppen.

(vi) Cytoplasma:
Cytoplasman är differentierad i en smal, yttre eller kortikal zon som kallas ektoplasma och en större, inre eller medullär region som kallas endoplasma.
(vii) Ektoplasma:
Ektoplasman (ectosarc eller cortex) är en permanent del av kroppen, påfallande avgränsad från endoplasman. Ektoplasman bildar ett fast, klart, tunt och tätt yttre skikt. Det innehåller trichocystor, cilier och fibrillära strukturer och avgränsas utvändigt av ett hölje som kallas pellicle.
(viii) Trikocystor:
Inbäddade i ektoplasman i rät vinkel mot ytan finns små spindelformade påsar som kallas trikocystor. En liten fläck på varje främre och bakre kant av sexhörningen markerar läget för en trikocysta. De är fyllda med en brytande, tät vätska som har en svällande substans, i den yttre änden finns ett koniskt huvud eller spik.
Trikocystorna ligger vinkelrätt i ektoplasman, de öppnas genom små porer på åsarna av de hexagonala områdena i pelliceln.
De uppstår från ciliernas kinetosomer, vandrar sedan och lokaliserar sig själva på lika långt avstånd i endoplasman. När djuret irriteras avges trichocystorna som långa klibbiga trådar. En utskjuten trikocysta har en ogenomskinlig spik, som liknar en omvänd spik, och ett långt randigt skaft, men skaftet syns inte i oskjutet tillstånd och bildas troligen under utskjutningen.
Trikocysternas funktion är osäker, men de avges som en reaktion på lokala kontakter och skador, de kan fungera som försvarsorganeller.
Men detta är osäkert eftersom trikocystorna är ineffektiva mot Didinium, det främsta rovdjuret på Paramecium, de kan vara till för att fixera djuret till en plats under födointag. Hos vissa ciliater fungerar trichocystorna som brottsorganeller. Efter att trichocystorna har avlossats regenereras de från kinetosomer.
Paramecium

(ix) Neuromotorium och tillhörande fibriller:
Enligt Lund (1933) finns på den vänstra dorsalväggen i cytopharynx ungefär i höjd med cytostomens bakre kant en mycket liten, tvåbent massa, neuromotorium. Från neuromotorium strålar fibriller ut i endoplasman.
Av dessa går vanligen fyra eller fler nästan till den dorsala kroppsväggen, men resten är kortare och inte bestämda i sitt läge. Alla benämns endoplasmatiska fibriller. Deras funktion är okänd, men de kan samordna de orala ciliernas matningsrörelser. Fibrillerna kan också ge mekaniskt stöd, elasticitet, kontraktilitet, ledningsförmåga och metabolisk påverkan.

(x) Endoplasma:
Endoplasman eller märgen är den mer flytande och voluminösa delen av cytoplasman som innehåller många cytoplasmiska granuler samt andra inneslutningar och strukturer av specialiserad karaktär. De cytoplasmatiska inneslutningarna är mitokondrier, Golgi-apparater, vakuoler, kristaller, granuler och kromidier osv. Andra strukturer, dvs. kärnor, kontraktila vakuoler och matvakuoler, finns också i endoplasman.
Kärnor:
I endoplasman nära cytostomen finns två kärnor, dvs. Paramecium är heterokaryotisk, en stor ellipsoid och granulär makronukleus och en annan liten kompakt mikronukleus.
Makronukleus är en iögonfallande, ellipsoidisk eller njurformad kropp. Den är av kompakt typ och innehåller fina trådar och tätt packade diskreta kromatingranuler av varierande storlek och inbäddade i en akromatisk matris. Den har många nukleoler och mycket mer kromatinmaterial (DNA).
Det är en somatisk eller vegetativ kärna. Den delar sig amitotiskt och kontrollerar djurets vegetativa funktioner (metaboliska aktiviteter). Den genomgår mitos.
Mikronkärnan är liten, kompakt och sfärisk. Den finns i allmänhet nära makronukleus ofta i en konkavitet. Fina kromatinkorn och kromatintrådar är jämnt fördelade i hela strukturen. Mikronukleusen delar sig mitotiskt och kontrollerar reproduktionen.
Antalet mikronukleus varierar med arten; det är ett hos P. caudatum, två hos P. aurelia och många hos P. multimicronucleatum. Mikronukleus innehåller en tydlig nukleolus hos P. aurelia men den finns inte hos P. caudatum. Moses (1949; 1950) har rapporterat att makronukleus och mikronukleus har samma kemiska sammansättning.
(xi) Kontraktila vakuoler:
Det finns två stora, vätskefyllda kontraktila vakuoler, var och en belägen nära kroppens ena ände nära dorsalytan. Deras position är fast (till skillnad från Amoeba), de ligger mellan ektoplasma och endoplasma, men de är tillfälliga organeller som försvinner med jämna mellanrum. Hos vissa arter verkar de ha ett fodermembran, i vilket fall de inte försvinner helt och hållet under systolen.
Med varje kontraktila vakuole är fem till tolv tubulära strålkanaler förbundna, som var och en består av en änddel, en lång ampulla som kollapsar när den är tom, och en kort injektionskanal som mynnar ut i vakuolen.

Kanalerna kommunicerar med en stor del av kroppen varifrån de tar upp vätskor och häller dem i vakuolen som på så sätt återskapas och växer i storlek, när den kontraktila vakuolen når sin maximala storlek drar den plötsligt ihop sig (systole) och släpper ut sitt innehåll genom en permanent por i pelliculan, därefter bildar kanalerna återigen de kontraktila vakuolerna, kanalerna försvinner inte helt eftersom de är permanenta strukturer.
De två kontraktila vakuolerna släpper ut oregelbundet, den bakre kontraherar snabbare eftersom den ligger nära cytopharynx och mer vatten kommer in i den. Kanalernas och de kontraktila vakuolernas huvudfunktion är hydrostatisk, de avlägsnar överskott av vatten från protoplasman, vattnet absorberas delvis och tas delvis in under födointag.
Den kvävehaltiga avfallssubstansen har ammoniakföreningar och en del urater som stöts ut från de kontraktila vakuolerna tillsammans med koldioxid, men det finns inga tecken på att utsöndringsämnena utsöndras av protoplasman i kanalerna eller de kontraktila vakuolerna.
Det finns en fast permanent cytopyge eller anus under peristomen på ena sidan av cytofarynx, genom vilken osmälta matrester och urater stöts ut.
Den elektronmikroskopiska studien av kontraktila vakuoler har avslöjat att varje kontraktila apparat består av några av endoplasmatiska retikulumets tubuli, nephridiala tubuli, matningskanaler, accessoriska vakuoler (radiella kanaler) och kontraktila huvudvakuoler. De accessoriska vakuolerna antas vara ampullerna till matningskanalerna (fig. 20.11).
(xii) Livsmedelsvakuoler:
Dessa är grovt sfäriska, icke-kontraktila kroppar som varierar i storlek och antal och som ligger i endoplasman. De innehåller intagna matpartiklar, huvudsakligen bakterier och en liten mängd vätska som avgränsas av ett tunt definierat membran. Volkonsky (1934) föreslog namnet gastrioler för dessa vakuoler. I anslutning till matvakuolerna finns de digestiva granulerna.
Förflyttning hos Paramecium Caudatum:
Paramecium Caudatum utför sin förflyttning med två metoder, nämligen metabolism eller kroppsförvrängningar och med hjälp av cilier.
(i) Metabolism eller kroppsrörelser:
Kroppen hos Paramecium caudatum är elastisk, den kan klämma sig igenom en passage som är smalare än kroppen, varefter kroppen återtar sin normala form. Denna tillfälliga förändring av kroppens form är metabolism, den åstadkoms hos Paramecium av protoplasman.
(ii) Ciliär förflyttning:
Lokomotion som åstadkoms av cilier är den huvudsakliga metoden. Cilierna kan slå framåt eller bakåt så att djuret kan simma framåt eller bakåt.
Normalt simmar djuret framåt och cilierna slår bakåt men snett, cilierna stelnar och böjer sig snabbt bakåt så att de nästan vidrör kroppsytan, detta kallas det effektiva slaget; sedan blir cilierna slappa och återgår långsamt till den ursprungliga vertikala positionen, detta kallas återhämtningsslaget.
Cilia i samma tvärgående rad slår tillsammans och de i samma längsgående rad slår en efter en från den främre till den bakre änden.

Denna samordnade rörelse av cilier kallas metakrona rytm, vilket beror på infraciliärsystemet; detta orsakar simning framåt av djuret. Men när kroppscilierna slår snett bakåt slår samtidigt de längre cilierna i munfåran kraftigare, vilket får den främre änden att svänga åt vänster.
Aktionen av kroppens och munspårets cilier gör att djuret roterar runt sin långa axel. Denna rotation är alltid åt vänster (utom hos P. calkinsi som roterar i en högerspiral).
Denna kombination av framåtgående rörelse, svängningar och rotation gör att djuret rör sig framåt i en spiralbana moturs. Denna bana har en rak axel, och samma kroppsyta hos djuret förblir mot spiralbanans axel. Men när de simmar bakåt roterar alla arter åt höger.
Ciliarslaget kan vändas så att cilierna rör sig snett framåt varigenom djuret simmar bakåt. Med hjälp av ciliärverkan rör sig Paramecium med en hastighet av 1500 mikrometer eller till och med mer per sekund.
Jennings hävdade att Parameceums spiralformade rörelse beror på att medan cilierna främst slår bakåt gör de det snett åt höger vilket gör att djuret rullar över till vänster.
Också denna svängning av kroppen mot den aborala ytan beror till stor del på den större kraften i det effektiva slaget av de orala cilierna som slår mer direkt bakåt. Resultatet – Parameciums rotation kring sin långa axel – gör det därigenom möjligt för Paramecium att följa en mer eller mindre rak kurs när det bildar stora spiraler.
Näring hos Paramecium Caudatum:
I Paramecium Caudatum är näringen holozoisk. Födan består huvudsakligen av bakterier och små protozoer. Paramecium väntar inte på maten utan jagar den aktivt.
Det påstås att Paramecium Caudatum visar ett val i valet av föda, men det verkar inte finnas någon grund för detta trots att den slukar endast vissa typer av bakterier; tillgängliga uppgifter tyder på att 2 till 5 miljoner individer av Bacillus coli slukas av ett enda Paramecium på 24 timmar. Det äter också encelliga växter som alger, kiselalger osv. och små bitar av djur och grönsaker.
Föda mekanism:
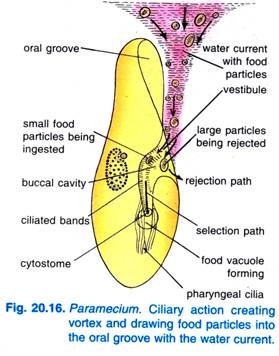
När Paramecium Caudatum kommer in i ett område med rikligt med föda kommer den till vila. Den äter endast i vila eller när den simmar mycket långsamt, den äter aldrig när den simmar snabbt. Slåendet av cilier i munfåran gör att en konisk virvel av födoladdat vatten sveps in i munfåran från ett avstånd före den främre änden (fig. 20.16).
Näringspartiklarna går sedan till vestibulum, varifrån vissa näringspartiklar stöts bort och kastas ut, men andra passerar in i cytostomen.
I slutet av cytopharynx bildas en matvakuol som fylls med matpartiklar. Quadrulus och peniculi kontrollerar matens passage till matvakuolen som bildas lateralt. När matvakuolen når en viss storlek griper de postbuckala fibrerna tag i matvakuolen och den kläms av av dem och börjar sin väg.
Vacuolen innehåller en del vatten förutom maten. Roterande strömmande rörelser av endoplasma, som kallas cyklos, för matvakuolerna längs en bestämd bana som funktionellt motsvarar en matsmältningskanal.
Kanalen börjar från cytofarynxens ände, sedan till den bakre sidan, sedan framåt för att cirkulera med endoplasman, sedan till den dorsala ytan, sedan mot den främre änden, sedan nedåt till cytopygen. I början av sin resa minskar matvakuolen i storlek för att sedan öka igen.

Förtäring och uppslutning hos Paramecium Caudatum:
Under cyklos sker förtäringen genom enzymer som utsöndras av protoplasman i vakuolerna. Vid matsmältningen omvandlas proteiner till aminosyror, kolhydrater till lösliga sockerarter och glykogen, och fetter smälts troligen också.
Innehållet i matens vakuoler är först surt (pH ca 4) och blir sedan alkaliskt, den huvudsakliga matsmältningen sker under den alkaliska fasen. Det osmälta materialet egestas genom cytopygen med viss kraft.
Cyklos kan demonstreras experimentellt; om mjölk färgad med Kongoröd matas till Paramecium, kommer mjölkens fettkulor i matvakuolerna först att bli röda på grund av enzymernas sura reaktion, sedan kommer de att skifta från lila till blått på grund av den alkaliska reaktionen, vakuolerna kommer att visa cyklosens förlopp.
Respiration och exkretion hos Paramecium Caudatum:
Gasutbytet (syre och koldioxid) sker genom den halvpermeabla hinnan liksom hos andra sötvattenprotozoer genom diffusionsprocessen. Paramecium Caudatum får sitt syre från det omgivande vattnet. Koldioxid och organiskt avfall som ammoniak från ämnesomsättningen utsöndras troligen genom att diffundera ut i vattnet i omvänd riktning.

6. Osmoregulering i Paramecium Caudatum:
Paramecium Caudatum har två kontraktila vakuoler, en främre och en bakre. De kontraktila vakuolernas funktion är osmoreglering, dvs. att reglera kroppens vatteninnehåll och kan även fungera vid utsöndring av kvävehaltiga avfallsprodukter som urea och ammoniak.
Överskott av vatten (på grund av kontinuerlig endosmosis) i cytoplasman utsöndras i endoplasmatiska retikulumets tubuli och går till nephridiala tubuli → matningskanaler → och samlas i ampulla i en serie av 6 till 11 strålande kanaler som konvergerar mot och mynnar ut i varje vakuole. Kanalerna är mest iögonfallande när en vacuole håller på att bildas.
När varje vacuole sväller (diastole) till en viss storlek drar den ihop sig (systole) och släpper ut till utsidan troligen genom en por. De kontraktila vakuolerna drar ihop sig växelvis med 10-20 sekunders mellanrum.
Den bakre kontraktila vakuolen arbetar snabbare än den främre vakuolen på grund av att cytofarynx tar in stora mängder vatten i den bakre regionen. De kontraktila vakuolerna upprätthåller en optimal koncentration av vatten i kroppens cytoplasma genom att göra sig av med överskottet.

Paramecium caudatums beteende:
Paramecium caudatums reaktioner på olika typer av stimuli lär man sig genom att studera dess reaktioner och grupperingen eller spridningen av individer i en kultur. Reaktionen är positiv om djuret rör sig mot ett stimuli och negativ om det rör sig bort. Vid en negativ stimulans fortsätter djuret att ge den undvikande reaktionen tills det flyr.
I en undvikande reaktion vänder ciliärslaget, djuret rör sig bakåt en kort sträcka och roterar sedan i en konisk bana genom att svänga den främre änden aboralt samtidigt som det svänger på den bakre spetsen. Alla justeringar görs genom försök och misstag. Försök har visat att den främre delen av djuret är känsligare än de andra delarna.
Parameciums reaktioner på olika stimuli kan grupperas enligt följande:
(i) Reaktioner på kontakt (Thigmotaxis):
Reaktionen på kontakt är varierande hos Paramecium. Om den främre änden berörs lätt med en fin spets uppstår en stark undvikande reaktion. När en simmande Paramecium kolliderar med något föremål i vattnet, men om den berörs någon annanstans kan ingen reaktion ske. En långsamt rörlig individ reagerar ofta, positivt på kontakt med ett föremål genom att komma till vila på det.
(ii) Reaktioner på kemikalier (Chemotaxis):
I allmänhet reagerar Paramecia på ett kemiskt stimuli genom att undvika reaktionen. Om en droppe svag saltlösning (0,5 procent) förs in i en Paramecium-population på en mikroslida, reagerar djuren med undvikande reaktion och ingen kommer in i droppen. För syror är dock reaktionen positiv även när koncentrationen är tillräckligt stark för att döda dem.
(iii) Reaktioner på temperatur (Thermotaxis):
Paramecium söker en optimal temperatur på 24 till 28 °C. När en temperaturförändring sker markant över eller under det optimala intervallet visar Paramecia en undvikande reaktion. Större värme stimulerar snabba rörelser och undvikande reaktioner tills djuren flyr eller dödas.
(iv) Reaktioner på ljus (Phototaxis):
Med undantag för den gröna Paramecium bursaria, som är positivt fototaktisk, är andra arter likgiltiga för vanligt ljus. När ljusintensiteten plötsligt och kraftigt ökas följer dock i allmänhet en negativ reaktion. Paramecia uppvisar en omedelbar negativ reaktion på ultravioletta strålar.
(v) Reaktioner på elektrisk ström (Galvanotaxis):
Paramecia reagerar på elektriska stimuli. När två elektroder placeras mitt emot varandra i en grund skål med Paramecia och en konstant ström tillförs, simmar alla organismer i samma riktning mot katoden eller den negativa elektroden där de koncentreras i stort antal.
Om riktningen på den elektriska strömmen vänds medan Paramecia simmar mot katoden, vänder organismerna riktning och simmar mot den nya katoden.

(vi) Reaktioner på vattenström (Rheotaxis):
Paramecia uppvisar en positiv reotaxi. I en svag vattenström rör sig Paramecia mestadels med strömmen med sina främre ändar uppströms.
(vii) Reaktioner på gravitation (Geotaxis):
Paramecia uppvisar i allmänhet en negativ reaktion på gravitation vilket ses i en kultur där många individer samlas tätt under ytfilmen med sina främre ändar överst. Om Paramecia introduceras i ett omvänt vattenfyllt U-format rör som stoppas i båda ändarna, rör de sig omedelbart uppåt i den horisontella delen av röret.
Reproduktion hos Paramecium Caudatam:
Paramecium Caudatum reproducerar sig asexuellt genom tvärgående binär fission och genomgår också flera typer av kärnreorganisering, såsom konjugering, endomixi, autogami, cytogami och hemixi osv.
(i) Tvärgående binär klyvning:
Transvers binär klyvning är den vanligaste typen av asexuell reproduktion hos Paramecium. Det är en klart unik asexuell process där ett fullvuxet exemplar delar sig i två dotterindivider utan att lämna ett föräldrakadaver.
Det plan i vilket delningen sker går genom cellens centrum och i ett plan i rät vinkel mot kroppens längdaxel. Delning av cellkroppen som helhet föregås alltid av delning av cellkärnorna; det verkar faktiskt som om reproduktionen inleds av kärnornas aktivitet och delning.

Paramecium caudatum förökar sig genom tvärgående binär klyvning under gynnsamma förhållanden. Vid binär fission delar sig mikrokärnan genom mitos i två dottermikronukleer, som förflyttar sig till motsatta ändar av cellen. Makronukleus förlängs och delas tvärgående genom amitos.
En annan cytopharynx knoppas av och två nya kontraktila vakuoler framträder, en nära främre änden och en annan nära bakre änden. Under tiden uppträder en konstriktionsfåra nära kroppens mitt och fördjupas tills cytoplasman är helt delad.
De resulterande två ”dotterparamecierna” är lika stora och innehåller var och en en uppsättning cellorganeller. Av de två producerade dotterparamecierna kallas den främre för proter och den bakre för opisthe. De växer till full storlek innan en ny delning sker.
Den binära klyvningsprocessen tar ungefär två timmar att genomföra och kan ske en till fyra gånger per dag, vilket ger 2 till 16 individer. Ungefär 600 generationer produceras under ett år.
Förökningshastigheten beror på de yttre förhållandena i form av föda, temperatur, kulturens ålder och befolkningstäthet; även på de inre faktorerna ärftlighet och fysiologi. Naturligtvis, om alla ättlingar till en individ skulle överleva och föröka sig, skulle antalet producerade paramecia snart motsvara jordens volym.
Tecknet klon används för att hänvisa till alla individer som har producerats från en individ genom klyvning. Alla medlemmar i en klon är ärftligt lika.
(ii) Konjugering:
Ordinariskt förökar sig Paramecium Caudatum genom binär klyvning under långa perioder, men med jämna mellanrum kan detta avbrytas genom att två djur förenas längs sina munytor för den sexuella processen konjugering.
Konjugation definieras som den tillfälliga föreningen av två individer som ömsesidigt utbyter mikrokärnmaterial. Det är en unik typ av en sexuell process där två organismer skiljs åt snart efter utbytet av kärnmaterial.
Sonneborn (1947) har på grundval av parningsbeteendet hos Paramecium Caudatum rapporterat att varje art av Paramecium existerar i ett antal sorter eller syngen. Vidare finns det inom varje syngen ett antal parningstyper vanligen två.
Parningstyperna förblir morfologiskt identiska men uppvisar fysiologiska skillnader. Hos P. aurelia finns det 14 syngen och 28 parningstyper, medan det hos P. caudatum finns 16 syngen och 32 parningstyper. Observationer har gjorts om att paramecia vanligtvis varken konjugerar med medlemmar av sin egen parningstyp eller med de andra sorterna, utan endast med den andra parningstypen av sin egen sort.

Faktorer som inducerar konjugering:
Faktorerna som inducerar konjugering varierar från art till art men några av dem anges nedan:
1. Konjugering sker vanligtvis under ogynnsamma levnadsförhållanden; svält eller brist på föda och särskild bakteriediet eller vissa kemikalier sägs inducera konjugeringsprocessen hos vissa arter av Paramecium.
2. Konjugationen sker efter cirka 300 asexuella generationer av binär klyvning, eller så alternerar den med binär klyvning med långa mellanrum för att föryngra den döende klonen, dvs. den sker hos de individer som måste ha genomgått ett önskvärt antal asexuella generationer, som sägs vara perioden av omognad, och sedan blir de könsmogna för att kunna konjugera.
3. Konjugering sker när det sker en förändring i parameciernas fysiologiska tillstånd, då sker den mellan sådana individer som är något mindre till storleken (210 mikrometer långa) och de befinner sig i ett skede som kan betraktas som en period av ohälsosam ålderdom; paramecierna i detta tillstånd kommer att dö om de inte tillåts konjugera.
4. Plötsligt mörker i ljusförhållanden och låga temperaturer sägs framkalla konjugationsprocessen hos vissa arter.
5. Konjugering sker inte under natt eller mörker; den börjar tidigt på morgonen och fortsätter till eftermiddagen.
6. Ett proteinhaltigt ämne i cilierna hos individer av parningstyp sägs inducera konjugationen.
Konjugationsprocessen:
Konjugationsprocessen skiljer sig åt hos olika arter av Paramecium, men nedanstående redogörelse gäller konjugationsprocessen hos P. caudatum (fig. 20.21).
Vid konjugering kommer två Paramecium caudatum (kallade prekonjuganter) av motsatta parningstyper av samma sort samman med sina ventrala ytor och förenas genom sina orala spår; deras cilier producerar en substans på kroppens yta som orsakar vidhäftning av de två konjugerande paramecia.
De slutar äta och deras munspårsapparat försvinner. Pelliculan och ektoplasma, vid kontaktpunkten, hos båda bryts ner, och en protoplasmatisk bro bildas mellan de två djuren. Nu kallas dessa individer för konjuganter.
I detta tillstånd simmar det konjugerande paret aktivt och samtidigt sker en rad kärnförändringar i varje konjugant som beskrivs nedan:
Makronukleus börjar sönderfalla, den blir lös i strukturen och bildar ett komplext tvinnat snår, under den senare halvan av konjugationsperioden kommer den slutligen att försvinna och absorberas i cytoplasman. Mikronukleusen i varje konjugant delar sig två gånger, varav den ena är en reduktionsdelning.
Sålunda produceras fyra haploida dottermikronukleer i varje konjugant. Tre av dessa fyra mikrokärnor degenererar i vardera, så att endast en återstår.
Den återstående mikronukleusen i varje konjugant delar sig mitotiskt i två ojämna pronukleer eller spelkärnor som bildar en större stationär kvinnlig prokärna och en mindre, aktiv migrerande manlig prokärna.
Den migrerande prokärnan hos den ena konjuganten korsar den protoplasmatiska bron och smälter samman med den stationära prokärnan hos den andra konjuganten för att bilda en synkaryon eller konjugationskärna där det diploida antalet kromosomer är återställt och det har skett ett utbyte av arvsmaterial.
Processen har jämförts med befruktning hos högre djur, men detta är inte befruktning eftersom inga könsceller är inblandade. Konjuganterna separeras nu (efter cirka 12-48 timmar) och kallas ex-konjuganter. Synkaryonet i varje ex-konjugant delar sig tre gånger för att bilda åtta mikrokärnor i varje ex-konjugant.
Fyra av de åtta mikronukleerna förstoras och blir makronukleer, och tre av de andra fyra mikronukleerna försvinner.
Den återstående mikronukleusen delar sig och samtidigt delar sig ex-konjuganten genom binär fission i två celler som vardera har två makronukleus och en mikronukleus. Cellerna och deras mikrokärnor delar sig en andra gång för att bilda fyra paramecia från varje ex-konjugant, så att var och en har en makronukleus och en mikronukleus.
Den nya makronukleusen, liksom även mikronukleusen, har tillverkats av nytt material. Dessa nya kärnor innehåller troligen en ny och annorlunda potential som återspeglas i de friska individerna.
Betydelse av konjugering:
En klon kommer att dö ut om det inte sker en nukleär omorganisation, men klonen kan föryngras så att den återfår sin tidigare styrka genom nukleär omorganisation, denna nukleära omorganisation åstadkoms genom konjugering, konjugering är alltså nödvändig för fortsatt binär klyvning.
Betydelsen av konjugering har sammanfattats nedan:
1. Konjugationen fungerar som en process av föryngring och omorganisering genom vilken rasens vitalitet återställs. Om konjugering inte sker under långa perioder försvagas paramecia och dör. (Woodruffs påstående om att hålla paramecia friska i 22 000 generationer utan konjugation motbevisas av Sonneborn, eftersom han visade att alla Woodruffs paramecia tillhörde samma parningstyp).
2. Det finns ingen könsskillnad hos konjuganterna även om endast paramecia av två olika parningstyper av samma sort kommer att konjugera.
3. Det finns ingen könsskillnad, men den aktiva migrerande prokärnan betraktas som hane och den stationära prokärnan som hona.
4. Konjugationen är endast en tillfällig förening, det sker ingen sammansmältning av cytoplasma och ingen zygot produceras, men kärnan hos varje ex-konjugant innehåller arvsmassa från två konjugerande individer.
5. Konjugationen medför att makronukleus ersätts med material från synkaryon, detta är en händelse av grundläggande betydelse. Vid binär klyvning fördelades makrokärnans kromosomer slumpmässigt på dottercellerna, fortsatt binär klyvning hade gjort klonen svag med vissa strukturella avvikelser.
Konjugationen åstadkommer bildandet av rätt antal kromosomer i makronukleus, så att rasen förnyas i styrka. Mikronukleusens roll är att återställa ett balanserat kromosom- och genkomplex.
Aberrant beteende vid reproduktion hos Paramecium Caudatum:
Paramecium Caudatum uppvisar vissa avvikelser i sitt kärnbeteende under fission och konjugation, dessa avvikelser är endomixis, autogamy, cytogamy och hemixis. Vid de tre första processerna sker genetisk rekombination och en ny makronukleus bildas från mikronukleus.
(i) Endomixis:
Woodruff och Erdmann (1914) rapporterade först en ny nukleär omorganisationsprocess, endomixis (Gr., endon = inom; mixis = blandning) i Paramecium aurelia, en bimikronukleär art (Fig. 20.22). Denna process beskrevs som en periodisk process där en ny makrokärnapparat produceras utan synkaryonbildning. Endomixis förekommer inom en enda individ.
Enligt Woodruff och Erdmann degenererar makrokärnan och mikronukleerna delar sig två gånger för att bilda åtta mikronukleer. Sex mikrokärnor degenererar och två återstår. När endast två mikrokärnor återstår delar sig djuret genom klyvning i två celler med vardera en mikrokärna.
Mikronukleus i varje dottercell delar sig två gånger för att bilda fyra mikronukleus. Två mikrokärnor förstoras och bildar makronukleer. Djuret och dess mikrokärnor delar sig så att det bildas två dotterindivider som var och en har en makronukleus och två mikrokärnor.
Endomixis förekommer i den sort av P. aurelia som inte konjugerar, därför kan effekten av endomixis vara densamma som av konjugationen eftersom båda processerna medför att makronukleus ersätts med material från mikronukleus, och båda processerna föryngrar rasens vitalitet.
Men de två processerna skiljer sig åt eftersom det inte sker någon sammansmältning av pronukleerna vid endomixis; endomixis kan jämföras med parthenogenes.
Hur som helst har vissa arbetare med goda skäl hävdat att endomixis inte är en giltig process och att den har beskrivits på grund av felaktiga observationer. Med all sannolikhet äger endomixis inte rum och det kan bara vara ett specialiserat fall av autogami.

Senare rapporterade Erdmamm och Woodruff (1916) endomixis hos Paramecium caudatum. Diller tror dock inte på giltigheten av denna process och anser att Erdmann och Woodruff helt enkelt har kombinerat stadierna hemixis och autogami till ett schema, endomixis.
(ii) Autogami:
Diller (1934, 1936) och Sonneborn (1950) beskrev en process av självbefruktning eller autogami som förekommer hos en enskild individ hos Paramecium aurelia (Fig. 20.23). Han rapporterade att vid autogami producerar tre mikrokärniga (pregamiska) delningar, som inbegriper mognad, de gametiska kärnorna (pronukleerna).
Under autogami hos P. aurelia delar sig de två mikronukleerna två gånger (en gång meiotiskt) för att bilda åtta mikronukleer, varav sex degenererar. Under tiden växer makronukleus till en skeinliknande massa som senare bryts i bitar för att absorberas i cytoplasman. Två av de åtta mikronukleerna, som pronukleer, går in i en protoplasmatisk kon som böljar ut nära cellmynningen.
De två pronukleerna smälter samman och bildar synkaryon. Synkaryonen delar sig två gånger och bildar fyra mikrokärnor. Två mikrokärnor blir makrokärnor. Parameciumet och dess mikrokärnor delar sig och bildar två dotterindivider, var och en med en makrokärna och två mikrokärnor. Denna process är avslutad på ungefär två dagar.
Autogami medför föryngring av rasen.
Det liknar konjugation såtillvida att den nya makrokärnan bildas av material från mikrokärnan, i den nya makrokärnan återställs det korrekta antalet kromosomer; och även i det faktum att fusion av två pronukleer sker. Men autogami skiljer sig från konjugation eftersom endast en individ deltar i autogami och den tillhandahåller båda pronukleerna, det är ett slags självbefruktning.
(iii) Hemixis:
Diller (1936) rapporterade hemixis hos Paramecium aurelia (Fig. 20.24). Hemixis är i första hand en process av fragmentering och delning av makrokärnor utan någon ovanlig mikrokärnaktivitet. Diller klassificerade hemixis i fyra typer, nämligen A, B, C och D enligt fig. 20.24 i P. aurelia, men han fann också alla typer i masskulturer av P. caudatum och P. multimicronucleatum.
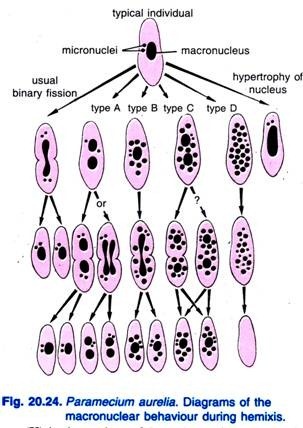
Typ A är den enklaste formen av hemixis som kännetecknas av en uppdelning av makronukleus i två eller flera delar. Denna delning är inte synkroniserad med mikrokärndelningen.
Typ B kännetecknas av att en till 20 eller fler kromatinkulor extruderas från makronukleus till cytoplasma.
Typ C kännetecknas av samtidig delning av makronukleus i två eller flera större delar och extrusion av makrokärnkulor till cytoplasma.
Typ D anses representera patologiska tillstånd där makrokärnan genomgår en fullständig fragmentering till kromatinkulor som slutligen försvinner från cellen. Mikronukleerna försvinner i allmänhet före upplösningen av makronukleerna.
(iv) Cytogami:
Wichterman (1939) har rapporterat en annan sexuell process i Paramecium caudatum, som han kallade cytogami. Vid cytogami sker inget utbyte av kärnor. I denna process kommer två individer samman genom sina ventrala ytor, men de två individernas pellicle bryts inte ner.
Mikronkärnan i varje individ delar sig tre gånger för att bilda åtta mikronkärnor, varav sex sönderfaller i varje individ. De två återstående mikrokärnorna smälter samman och bildar en synkaryon i varje cell. Djuren skiljer sig nu åt.
Cytogami skiljer sig från autogami genom att det finns två djur i kontakt med varandra, men det liknar autogami och konjugation genom sammansmältningen av två pronukleier. Cytogami skiljer sig från konjugation genom att det inte sker något kärnutbyte mellan de två djur som möts.
Vissa cytoplasmatiska partiklar som rapporterats i Paramecium caudatum:
(i) Kappapartiklar:
1938 rapporterade T.M. Sonneborn att vissa raser (kända som mördare eller mördarstam) av Paramecium producerar ett giftigt ämne, kallat paramecin, som är dödligt för andra individer som kallas känsliga. Paramecinet är vattenlösligt och diffust och är för sin produktion beroende av vissa partiklar som finns i cytoplasman hos Paramecium (mördarstammen).
Dessa partiklar kallas kappa-partiklar. Kappa-partiklarna har DNA och RNA. Ett mördarparamecium kan innehålla hundratals kappa-partiklar. Den detaljerade studien av dessa partiklar har visat att en dominant gen (K) i Parameciums kärna är nödvändig för att kappa-partiklar ska kunna existera, föröka sig och producera paramecin.
(ii) mµ, partiklar:
R.W. Siegel (1952) rapporterade en annan typ av mördarpartiklar i cytoplasman hos vissa Paramecium. Ett Paramecium med mµ-partiklar kallas mate killer eftersom när det konjugerar med ett Paramecium utan några mµ-partiklar som kallas mate sensitive, så dödar det det senare. MP-partiklarna består också av DNA, RNA osv.
Dessa partiklar finns endast i de paramecia vars mikronukleus innehåller minst en dominant gen av något av två par av obundna kromosomala gener (M1 och M2).
(iii) Pi-partiklar:
Dessa partiklar antas vara den muterade formen av kappa-partiklar, men de producerar inte någon typ av giftigt ämne.
(iv) Lambdapartiklar:
Dessa partiklar rapporteras i killer paramecia och sägs producera något ämne som är ansvarigt för att orsaka lysis eller sönderdelning av känsliga paramecia, dvs. som inte har det.