斜頸という言葉は、手足の指骨や中手骨・中足骨を含む骨形成異常のグループを包含しています。 5つのタイプ(A~E)といくつかのサブタイプ(A1~A4; E1~E3)があります。 常染色体優性遺伝をし、浸透度は様々である。 C型は第2指、第3指、第5指の中指と第1中手骨の短縮が特徴で、人差し指の尺側偏位、多指症、第2指と第3指の近位または中指の顕著な過分節がみられることもある。 1-3 「天使形」指骨(図1A)は、BDCに特徴的ではあるが、天使形指骨形成不全と呼ばれる疾患でも見られるため、予後不良ではない。 1 BDCでは、この特徴は手の骨の骨片閉鎖が完了すると正常化し、単純性腕足長となります。
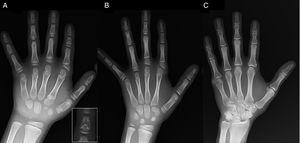
(A) プロブランドの左手のX線写真(年代7歳、骨年齢5歳)、異常な第1中手(近位と遠位の2枚の骨端板)と第2、3、5指の中指の短縮を認める。 第2指は尺側偏位を示し、第4指は最も影響が少なく、左手で最も長い。 第2指と第3指の近位骨端は形成不全で、第2指では天使型の中指骨が目立ちます(A の挿入部分)。 (B). プロバンドの妹(5歳半、骨年齢の遅れはない)の左手のX線写真で、第2、3、5指の中指骨は短縮しており、第4指は正常である。 この場合、第一中手骨は正常であった。 最も顕著な特徴は、第2指の近位骨端の形状が兄と同様に三角形であることと、中指の指骨の形状が台形であることである。 兄と同様に第2指は尺側偏位を呈している。 (C)プロバンドの父親の左手のX線写真。小児期に手術で矯正された軸後六本指の骨残骸のみが認められる。
低身長(z-score, -1.8)の評価で紹介されたBDCに適合する放射線学的特徴を有する7歳男児の症例を報告する。 父親は片側性軸後多指症を有し,父方の叔父にも両側性多指症がみられた。 左手と手首のX線検査では、骨年齢が年齢より2歳若く、異常が認められたため、骨格検査を実施した。 その結果、手に異常(Fig.1A)、足に軽微な異常(一部の足指近位指骨に軽度の骨端異形成)が検出された。 6歳の妹のX線検査では、手指の病変はより顕著ではないが、本人と類似していた(Fig.1B)。 父親の手のX線写真(Fig.1C)では,小児期に手術で矯正された後軸性六本指の骨の残骸が確認されたのみであった
GDF5遺伝子の配列決定により,候補者のエクソン2におけるヘテロ接合性の点置換(c.1462A>T)。その結果、早発停止コドン(p.Lys488*、ナンセンス変異)が生じ、野生タンパク質より14アミノ酸短い切断型タンパク質となった(図2A)。 この変異は患者の父と姉にも見られたが、健康な母には見られなかった(Fig.2B)。 Fig. 2Cはこの家族の血統を示したものである。 これはおそらく病原性で常染色体優性遺伝のパターンを持つ新規変異である。
GDF5遺伝子に検出された新規の変異。 (A)プロバンドのエクソン2の遺伝子配列。早発停止コドンと切断タンパク質(p.Lys488*)をもたらすc.1462A>T変異を示す(我々は停止コドンの正常位置を緑の四角で表示した)。 (B)健常な母親の同領域の正常な配列。 (C) この家族の血統図:黒字の父親と子供二人はGDF5の変異が確認されている。 健常な母親は白色で表示されている。 灰色の四角は両軸後多指症を持つ父方の叔父で、おそらくこの変異を持つ。
Growth Differentiation factor 5 (GDF5) は骨形成タンパク質と密接に関連し、トランスフォーミング成長因子βスーパーファミリーに属し、胚の骨格と関節の発達に関与している4。 GDF5遺伝子は、骨格奇形を伴う疾患の変異のホットスポットである5。ほとんどのホモ接合体または複合ヘテロ接合体の変異は、重篤な疾患と関連している。 ホモ接合型あるいは複合型ヘテロ接合型変異の多くは、Grebe型軟骨異形成症(OMIM 200700)、Hunter-Thomson型骨端症(OMIM 201250)あるいはDu Pan症候群(OMIM 228900)などの重篤な疾患と関連している。 一方、ヘテロ接合型変異は、より軽度の骨格形成異常と関連している。近位指骨症1 B (OMIM 615298) と多発性指骨症症候群2型 (OMIM 610017) は、いずれもミスセンス変異で機能獲得を伴い、腕足指症A1およびA2型もミスセンス変異であるが機能喪失を伴う。5 C型は機能低下を伴うヘテロ接合型変異を伴うが、劣性遺伝をする症例も3例報告されている6。 BDCに関連する変異の多くは、遺伝子のプロドメイン部分のフレームシフト変異であり、成熟ドメインの変異の多くはミスセンス変異で、表現型は非常に多様である。4
今回紹介する家族は、タンパク質の活性ドメインをコードする領域にナンセンス変異があり、最後の14アミノ酸が除去された状態となっている。 最初の変異は、今回紹介するファミリーで変異したアミノ酸の直前のアミノ酸に同様の変異(p.Tyr487*/c.1461T>G)から、両者とも変異型モノマーを生じ、GDF5の機能的ハプロインフィシェンシーを引き起こし、BDCを引き起こすことが示唆された。
資金提供
本研究は、スペイン政府の2013-2016 R&D&I計画に組み込まれ、Instituto de Salud Carlos III(ISCIII)の研究評価・推進副総局が共同出資したPI13/00467およびPI13/01295プロジェクトにより一部資金提供されたものです。 欧州地域開発基金(ERDF)およびCentro de Investigación Biomédica en Red sobre Fisiopatología de la Obesidad y la Nutrición (Biomedical Research Networking Centre on the Physiopathology of Obesity and Nutrition ), ISCIII, Madrid.